Eye
From Wikipedia, the free encyclopedia
| Eye | |
|---|---|
 |
|
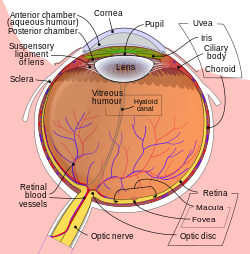
| Schematic diagram of the vertebrate eye. | |
 |
|
| Compound eye of Antarctic krill |
Eyes are organs that detect light, and send signals along the optic nerve to the visual and other areas of the brain[citation needed]. Complex optical systems with resolving power have come in ten fundamentally different forms, and 96% of animal species possess a complex optical system.[1] Image-resolving eyes are present in cnidaria, mollusks, chordates, annelids and arthropods.[2]
The simplest "eyes", in even unicellular organisms, do nothing but detect whether the surroundings are light or dark, which is sufficient for the entrainment of circadian rhythms. From more complex eyes, retinal photosensitive ganglion cells send signals along the retinohypothalamic tract to the suprachiasmatic nuclei to effect circadian adjustment.
Contents |
[edit] Overview

Complex eyes can distinguish shapes and colors. The visual fields of many organisms, especially predators, involve large areas of binocular vision to improve depth perception; in other organisms, eyes are located so as to maximise the field of view, such as in rabbits and horses.
The first proto-eyes evolved among animals 540 million years ago, about the time of the so-called Cambrian explosion.[citation needed] The last common ancestor of animals possessed the biochemical toolkit necessary for vision, and more advanced eyes have evolved in 96% of animal species in 6 of the thirty-something[note 1] main phyla.[1] In most vertebrates and some mollusks, the eye works by allowing light to enter it and project onto a light-sensitive panel of cells, known as the retina, at the rear of the eye. The cone cells (for color) and the rod cells (for low-light contrasts) in the retina detect and convert light into neural signals for vision. The visual signals are then transmitted to the brain via the optic nerve. Such eyes are typically roughly spherical, filled with a transparent gel-like substance called the vitreous humour, with a focusing lens and often an iris; the relaxing or tightening of the muscles around the iris change the size of the pupil, thereby regulating the amount of light that enters the eye,[3] and reducing aberrations when there is enough light.[4]
The eyes of cephalopods, fish, amphibians and snakes usually have fixed lens shapes, and focusing vision is achieved by telescoping the lens — similar to how a camera focuses.[5]
Compound eyes are found among the arthropods and are composed of many simple facets which, depending on the details of anatomy, may give either a single pixelated image or multiple images, per eye. Each sensor has its own lens and photosensitive cell(s). Some eyes have up to 28,000 such sensors, which are arranged hexagonally, and which can give a full 360-degree field of vision. Compound eyes are very sensitive to motion. Some arthropods, including many Strepsiptera, have compound eyes of only a few facets, each with a retina capable of creating an image, creating vision. With each eye viewing a different thing, a fused image from all the eyes is produced in the brain, providing very different, high-resolution images.
Possessing detailed hyperspectral color vision, the Mantis shrimp has been reported to have the world's most complex color vision system.[6] Trilobites, which are now extinct, had unique compound eyes. They used clear calcite crystals to form the lenses of their eyes. In this, they differ from most other arthropods, which have soft eyes. The number of lenses in such an eye varied, however: some trilobites had only one, and some had thousands of lenses in one eye.
In contrast to compound eyes, simple eyes are those that have a single lens. For example, jumping spiders have a large pair of simple eyes with a narrow field of view, supported by an array of other, smaller eyes for peripheral vision. Some insect larvae, like caterpillars, have a different type of simple eye (stemmata) which gives a rough image. Some of the simplest eyes, called ocelli, can be found in animals like some of the snails, which cannot actually "see" in the normal sense. They do have photosensitive cells, but no lens and no other means of projecting an image onto these cells. They can distinguish between light and dark, but no more. This enables snails to keep out of direct sunlight. In organisms dwelling near deep-sea vents, compound eyes have been secondarily simplified and adapted to spot the infra-red light produced by the hot vents - in this way the bearers can spot hot springs and avoid being boiled alive.[7]
[edit] Evolution
| This section requires expansion. |
Visual pigments appear to have a common ancestor and were probably involved in circadian rhythms or reproductive timing in simple organisms. Complex vision - associated with dedicated visual organs, or eyes - evolved many times in different lineages.
[edit] Types of eye
Nature has produced ten different eye layouts — indeed every way of capturing an image has evolved at least once in nature, with the exception of zoom and Fresnel lenses. Eye types can be categorized into "simple eyes", with one concave chamber, and "compound eyes", which comprise a number of individual lenses laid out on a convex surface.[1] Note that "simple" does not imply a reduced level of complexity or acuity. Indeed, any eye type can be adapted for almost any behaviour or environment. The only limitations specific to eye types are that of resolution — the physics of compound eyes prevents them from achieving a resolution better than 1°. Also, superposition eyes can achieve greater sensitivity than apposition eyes, so are better suited to dark-dwelling creatures.[1] Eyes also fall into two groups on the basis of their photoreceptor's cellular construction, with the photoreceptor cells either being cilliated (as in the vertebrates) or rhabdomic. These two groups are not monophyletic; the cnidaira also possess cilliated cells, [8] and some annelids possess both.[9]
[edit] Simple eyes
[edit] Pit eyes
Pit eyes, also known as stemma, are eye-spots which may be set into a pit to reduce the angles of light that enters and affects the eyespot, to allow the organism to deduce the angle of incoming light.[1] Found in about 85% of phyla, these basic forms were probably the precursors to more advanced types of "simple eye". They are small, comprising up to about 100 cells covering about 100 µm.[1] The directionality can be improved by reducing the size of the aperture, by incorporating a reflective layer behind the receptor cells, or by filling the pit with a refractile material.[1]
[edit] Pinhole eye

The pinhole eye is an "advanced" form of pit eye incorporating these improvements, most notably a small aperture (which may be adjustable) and deep pit. It is only found in the nautiloids.[1] Without a lens to focus the image, it produces a blurry image, and will blur out a point to the size of the aperture. Consequently, nautiloids can't discriminate between objects with an angular separation of less than 11°.[1] Shrinking the aperture would produce a sharper image, but let in less light.[1]
[edit] Spherical lensed eye
The resolution of pit eyes can be greatly improved by incorporating a material with a higher refractive index to form a lens, which may greatly reduce the blur radius encountered — hence increasing the resolution obtainable.[1] The most basic form, still seen in some gastropods and annelids, consists of a lens of one refractive index. A far sharper image can be obtained using materials with a high refractive index, decreasing to the edges — this decreases the focal length and thus allows a sharp image to form on the retina.[1] This also allows a larger aperture for a given sharpness of image, allowing more light to enter the lens; and a flatter lens, reducing spherical aberration.[1] Such an inhomogeneous lens is necessary in order for the focal length to drop from about 4 times the lens radius, to 2.5 radii.[1]
Heterogeneous eyes have evolved at least eight times — four or more times in gastropods, once in the copepods, once in the annelids and once in the cephalopods.[1] No aquatic organisms possess homogeneous lenses; presumably the evolutionary pressure for a heterogeneous lens is great enough for this stage to be quickly "outgrown".[1]
This eye creates an image that is sharp enough that motion of the eye can cause significant blurring. To minimize the effect of eye motion while the animal moves, most such eyes have stabilizing eye muscles.[1]
The ocelli of insects bear a simple lens, but their focal point always lies behind the retina; consequently they can never form a sharp image. This capitulates the function of the eye. Ocelli (pit-type eyes of arthropods) blur the image across the whole retina, and are consequently excellent at responding to rapid changes in light intensity across the whole visual field — this fast response is further accelerated by the large nerve bundles which rush the information to the brain.[10] Focussing the image would also cause the sun's image to be focussed on a few receptors, with the possibility of damage under the intense light; shielding the receptors would block out some light and thus reduce their sensitivity.[10] This fast response has led to suggestions that the ocelli of insects are used mainly in flight, because they can be used to detect sudden changes in which way is up (because light, especially UV light which is absorbed by vegetation, usually comes from above).[10]
[edit] Weaknesses
One weakness of this eye construction is that chromatic aberration is still quite high[1] — although for organisms without color vision, this is a very minor concern.
A weakness of the vertebrate eye is the blind spot which results from a gap in the retina where the optic nerve exits at the back of the eye; the cephalopod eye has no blind spot as the retina is in the opposite orientation.
[edit] Multiple lenses
Some marine organisms bear more than one lens; for instance the copeopod Pontella has three. The outer has a parabolic surface, countering the effects of spherical aberration while allowing a sharp image to be formed. Copilla's eyes have two lenses, which move in and out like a telescope.[1] Such arrangements are rare and poorly understood, but represent an interesting alternative construction. An interesting use of multiple lenses is seen in some hunters such as eagles and jumping spiders, which have a refractive cornea (discussed next): these have a negative lens, enlarging the observed image by up to 50% over the receptor cells, thus increasing their optical resolution.[1]
[edit] Refractive cornea
In the eyes of most terrestrial vertebrates (along with spiders and some insect larvae) the vitreous fluid has a higher refractive index than the air, relieving the lens of the function of reducing the focal length. This has freed it up for fine adjustments of focus, allowing a very high resolution to be obtained.[1] As with spherical lenses, the problem of spherical aberration caused by the lens can be countered either by using an inhomogeneous lens material, or by flattening the lens.[1] Flattening the lens has a disadvantage: the quality of vision is diminished away from the main line of focus, meaning that animals requiring all-round vision are detrimented. Such animals often display an inhomogeneous lens instead.[1]
As mentioned above, a refractive cornea is only useful out of water; in water, there is no difference in refractive index between the vitreous fluid and the surrounding water. Hence creatures which have returned to the water — penguins and seals, for example — lose their refractive cornea and return to lens-based vision. An alternative solution, borne by some divers, is to have a very strong cornea.[1]
[edit] Reflector eyes
An alternative to a lens is to line the inside of the eye with " mirrors", and reflect the image to focus at a central point.[1] The nature of these eyes means that if one were to peer into the pupil of an eye, one would see the same image that the organism would see, reflected back out.[1]
Many small organisms such as rotifers, copeopods and platyhelminths use such organs, but these are too small to produce usable images.[1] Some larger organisms, such as scallops, also use reflector eyes. The scallop Pecten has up to 100 millimeter-scale reflector eyes fringing the edge of its shell. It detects moving objects as they pass successive lenses.[1]
There is at least one vertebrate, the spookfish, whose eyes include reflective optics for focusing of light. Each of the two eyes of a spookfish collects light from both above and below; the light coming from the above is focused by a lens, while that coming from below, by a curved mirror composed of many layers of small reflective plates made of guanine crystals.[11]
[edit] Compound eyes
A compound eye may consist of thousands of individual photoreception units. The image perceived is a combination of inputs from the numerous ommatidia (individual "eye units"), which are located on a convex surface, thus pointing in slightly different directions. Compared with simple eyes, compound eyes possess a very large view angle, and can detect fast movement and, in some cases, the polarization of light.[12] Because the individual lenses are so small, the effects of diffraction impose a limit on the possible resolution that can be obtained. This can only be countered by increasing lens size and number — to see with a resolution comparable to our simple eyes, humans would require compound eyes which would each reach the size of their head.
Compound eyes fall into two groups: apposition eyes, which form multiple inverted images, and superposition eyes, which form a single erect image.[13] Compound eyes are common in arthropods, and are also present in annelids and some bivalved molluscs.[14]
Compound eyes, in arthropods at least, grow at their margins by the addition of new ommatidia.[15]
[edit] Apposition eyes
Apposition eyes are the most common form of eye, and are presumably the ancestral form of compound eye. They are found in all arthropod groups, although they may have evolved more than once within this phylum.[1] Some annelids and bivalves also have apposition eyes. They are also possessed by Limulus, the horseshoe crab, and there are suggestions that other chelicerates developed their simple eyes by reduction from a compound starting point.[1] (Some caterpillars appear to have evolved compound eyes from simple eyes in the opposite fashion.)
Apposition eyes work by gathering a number of images, one from each eye, and combining them in the brain, with each eye typically contributing a single point of information.
The typical apposition eye has a lens focusing light from one direction on the rhabdom, while light from other directions is absorbed by the dark wall of the ommatidium. In the other kind of apposition eye, found in the Strepsiptera, lenses are not fused to one another, and each forms an entire image; these images are combined in the brain. This is called the schizochroal compound eye or the neural superposition eye. Because images are combined additively, this arrangement allows vision under lower light levels.[1]
[edit] Superposition eyes
The second type is named the superposition eye. The superposition eye is divided into three types; the refracting, the reflecting and the parabolic superposition eye. The refracting superposition eye has a gap between the lens and the rhabdom, and no side wall. Each lens takes light at an angle to its axis and reflects it to the same angle on the other side. The result is an image at half the radius of the eye, which is where the tips of the rhabdoms are. This kind is used mostly by nocturnal insects. In the parabolic superposition compound eye type, seen in arthropods such as mayflies, the parabolic surfaces of the inside of each facet focus light from a reflector to a sensor array. Long-bodied decapod crustaceans such as shrimp, prawns, crayfish and lobsters are alone in having reflecting superposition eyes, which also has a transparent gap but uses corner mirrors instead of lenses.
[edit] Parabolic superposition
This eye type functions by refracting light, then using a parabolic mirror to focus the image; it combines features of superposition and apposition eyes.[7]
[edit] Other

Good fliers like flies or honey bees, or prey-catching insects like praying mantis or dragonflies, have specialized zones of ommatidia organized into a fovea area which gives acute vision. In the acute zone the eye are flattened and the facets larger. The flattening allows more ommatidia to receive light from a spot and therefore higher resolution.
There are some exceptions from the types mentioned above. Some insects have a so-called single lens compound eye, a transitional type which is something between a superposition type of the multi-lens compound eye and the single lens eye found in animals with simple eyes. Then there is the mysid shrimp Dioptromysis paucispinosa. The shrimp has an eye of the refracting superposition type, in the rear behind this in each eye there is a single large facet that is three times in diameter the others in the eye and behind this is an enlarged crystalline cone. This projects an upright image on a specialized retina. The resulting eye is a mixture of a simple eye within a compound eye.
Another version is the pseudofaceted eye, as seen in Scutigera. This type of eye consists of a cluster of numerous ocelli on each side of the head, organized in a way that resembles a true compound eye.
The body of Ophiocoma wendtii, a type of brittle star, is covered with ommatidia, turning its whole skin into a compound eye. The same is true of many chitons.
[edit] Relationship to lifestyle
Eyes are generally adapted to the environment and lifestyle of the organism which bears them. For instance, the distribution of photoreceptors tends to match the area in which the highest acuity is required, with horizon-scanning organisms, such as those that live on the African plains, having a horizontal line of high-density ganglia, while tree-dwelling creatures which require good all-round vision tend to have a symmetrical distribution of ganglia, with acuity decreasing outwards from the centre.
Of course, for most eye types, it is impossible to diverge from a spherical form, so only the density of optical receptors can be altered. In organisms with compound eyes, it is the number of ommatidia rather than ganglia that reflects the region of highest data acquisition.[1] Optical superposition eyes are constrained to a spherical shape, but other forms of compound eyes may deform to a shape where more ommatidia are aligned to, say, the horizon, without altering the size or density of individual ommatidia.[16] Eyes of horizon-scanning organisms have stalks so they can be easily aligned to the horizon when this is inclined, for example if the animal is on a slope.[17] An extension of this concept is that the eyes of predators typically have a zone of very acute vision at their centre, to assist in the identification of prey.[16] In deep water organisms, it may not be the centre of the eye that is enlarged. The hyperiid amphipods are deep water animals that feed on organisms above them. Their eyes are almost divided into two, with the upper region thought to be involved in detecting the silhouettes of potential prey — or predators — against the faint light of the sky above. Accordingly, deeper water hyperiids, where the light against which the silhouettes must be compared is dimmer, have larger "upper-eyes", and may lose the lower portion of their eyes altogether.[16] Depth perception can be enhanced by having eyes which are enlarged in one direction; distorting the eye slightly allows the distance to the object to be estimated with a high degree of accuracy.[7]
Acuity is higher among male organisms that mate in mid-air, as they need to be able to spot and assess potential mates against a very large backdrop.[16] On the other hand, the eyes of organisms which operate in low light levels, such as around dawn and dusk or in deep water, tend to be larger to increase the amount of light that can be captured.[16]
It is not only the shape of the eye that may be affected by lifestyle. Eyes can be the most visible parts of organisms, and this can act as a pressure on organisms to have more transparent eyes at the cost of function.[16]
Eyes may be mounted on stalks to provide better all-round vision, by lifting them above an organism's carapace; this also allows them to track predators or prey without moving the head.[7]
[edit] Acuity

Visual acuity is often measured in cycles per degree (CPD), which measures an angular resolution, or how much an eye can differentiate one object from another in terms of visual angles. Resolution in CPD can be measured by bar charts of different numbers of white — black stripe cycles. For example, if each pattern is 1.75 cm wide and is placed at 1 m distance from the eye, it will subtend an angle of 1 degree, so the number of white — black bar pairs on the pattern will be a measure of the cycles per degree of that pattern. The highest such number that the eye can resolve as stripes, or distinguish from a gray block, is then the measurement of visual acuity of the eye.
For a human eye with excellent acuity, the maximum theoretical resolution would be 50 CPD[18] (1.2 arcminute per line pair, or a 0.35 mm line pair, at 1 m). A rat can resolve only about 1 to 2 CPD.[19] A horse has higher acuity through most of the visual field of its eyes than a human has, but does not match the high acuity of the human eye's central fovea region.
Spherical aberration limits the resolution of a 7 mm pupil to about 3 arcminutes per line pair. At a pupil diameter of 3 mm, the spherical aberration is greatly reduced, resulting in an improved resolution of approximately 1.7 arcminutes per line pair.[20] A resolution of 2 arcminutes per line pair, equivalent to a 1 arcminute gap in an optotype, corresponds to 20/20 (normal vision) in humans.
[edit] Color
All organisms are restricted to a small range of the electromagnetic spectrum; this varies from creature to creature, but is mainly between 400 and 700 nm[21]. This is a rather small section of the electromagnetic spectrum, probably reflecting the submarine evolution of the organ: water blocks out all but two small windows of the EM spectrum, and there has been no evolutionary pressure among land animals to broaden this range.[22]
The most sensitive pigment, rhodopsin, has a peak response at 500 nm.[23] Small changes to the genes coding for this protein can tweak the peak response by a few nm;[2] pigments in the lens can also "filter" incoming light, changing the peak response.[2] Many[quantify] organisms are unable to discriminate between colors, seeing instead in shades of "grey"; color vision necessitates a range of pigment cells which are primarily sensitive to smaller ranges of the spectrum. In primates, geckos, and other organisms, these take the form of cone cells, from which the more sensitive rod cells evolved[verification needed].[23] Even if organisms are physically capable of discriminating different colors, this does not necessarily mean that they can perceive the different colors; only with behavioral tests can this be deduced.[2]
Most organisms with color vision are able to detect ultraviolet light. This high energy light can be damaging to receptor cells. With a few exceptions (snakes, placental mammals), most organisms avoid these effects by having absorbent oil droplets around their cone cells. The alternative, developed by organisms that had lost these oil droplets in the course of evolution, is to make the lens impervious to UV light — this precludes the possibility of any UV light being detected, as it does not even reach the retina.[23]
[edit] Rods and cones
The retina contains two major types of light-sensitive photoreceptor cells used for vision: the rods and the cones.
Rods cannot distinguish colors, but are responsible for low-light black-and-white (scotopic) vision; they work well in dim light as they contain a pigment, visual purple, which is sensitive at low light intensity, but saturates at higher (photopic) intensities. Rods are distributed throughout the retina but there are none at the fovea and none at the blind spot. Rod density is greater in the peripheral retina than in the central retina.
Cones are responsible for color vision. They require brighter light to function than rods require. There are three types of cones, maximally sensitive to long-wavelength, medium-wavelength, and short-wavelength light (often referred to as red, green, and blue, respectively, though the sensitivity peaks are not actually at these colors). The color seen is the combined effect of stimuli to, and responses from, these three types of cone cells. Cones are mostly concentrated in and near the fovea. Only a few are present at the sides of the retina. Objects are seen most sharply in focus when their images fall on this spot, as when one looks at an object directly. Cone cells and rods are connected through intermediate cells in the retina to nerve fibers of the optic nerve. When rods and cones are stimulated by light, the nerves send off impulses through these fibers to the brain.[23]
[edit] Pigment
The pigment molecules used in the eye are various, but can be used to define the evolutionary distance between different groups, and can also be an aid in determining which are closely related – although problems of convergence do exist.[23]
Opsins are the pigments involved in photoreception. Other pigments, such as melanin, are used to shield the photoreceptor cells from light leaking in from the sides. The opsin protein group evolved long before the last common ancestor of animals, and has continued to diversify since.[2]
There are two types of opsin involved in vision; c-opsins, which are associated with ciliary-type photoreceptor cells, and r-opsins, associated with rhabdomeric photoreceptor cells.[24] The eyes of vertebrates usually contain cilliary cells with c-opsins, and (bilaterian) invertebrates have rhabdomeric cells in the eye with r-opsins. However, some ganglion cells of vertebrates express r-opsins, suggesting that their ancestors used this pigment in vision, and that remnants survive in the eyes.[24] Likewise, c-opsins have been found to be expressed in the brain of some invertebrates. They may have been expressed in ciliary cells of larval eyes, which were subsequently resorbed into the brain on metamorphosis to the adult form.[24] C-opsins are also found in some derived bilaterian-invertebrate eyes, such as the pallial eyes of the bivalve molluscs; however, the lateral eyes (which were presumably the ancestral type for this group, if eyes evolved once there) always use r-opsins.[24] Cnidaria, which are an outgroup to the taxa mentioned above, express c-opsins - but r-opsins are yet to be found in this group.[24] Incidentally, the melanin produced in the cnidaria is produced in the same fashion as that in vertebrates, suggesting the common descent of this pigment.[24]
[edit] See also
[edit] References
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af Land, M F; Fernald, R D (1992). "The Evolution of Eyes". Annual Review of Neuroscience 15: 1–29. doi:.
- ^ a b c d e Frentiu, Francesca D.; Adriana D. Briscoe (2008), "A butterfly eye's view of birds", BioEssays 30: 1151, doi:
- ^ Nairne, James (2005). Psychology. Belmont: Wadsworth Publishing. ISBN 049503150x. OCLC 61361417. http://books.google.com/books?id=6MqkLT-Q0oUC&pg=PA146&dq=iris+intitle:psychology+inauthor:Nairne&lr=&as_brr=0&ei=jD_dR4jcDorysgP6yMXqAQ&sig=225aBFAHEbzLuL9jBuNMCCFDdHw.
- ^ Vicki Bruce, Patrick R. Green, and Mark A. Georgeson (1996). Visual Perception: Physiology, Psychology and Ecology. Psychology Press. pp. 20. ISBN 0863774504. http://books.google.com/books?id=ukvei0wge_8C&pg=PA20&dq=iris+aberrations+intitle:psychology&lr=&as_brr=0&ei=EEDdR-2gAZOMtAPWpJX4AQ&sig=lTs0jcmjOgcd_P0-lo24zbrsfio.
- ^ BioMedia Associates Educational Biology Site: What animal has a more sophisticated eye, Octopus or Insect?
- ^ Who You Callin' "Shrimp"? – National Wildlife Magazine
- ^ a b c d Cronin, Thomas W. (2008). "Exceptional Variation on a Common Theme: the Evolution of Crustacean Compound Eyes". Evolution Education and Outreach 1: 463. doi:.
- ^ Kozmik, Zbynek; Ruzickova, Jana; Jonasova, Kristyna; Matsumoto, Yoshifumi; Vopalensky, Pavel; Kozmikova, Iryna; Strnad, Hynek; Kawamura, Shoji; Piatigorsky, Joram; Paces, Vaclav; Vlcek, Cestmir (2008), "Assembly of the cnidarian camera-type eye from vertebrate-like components" (PDF), Proceedings of the National Academy of Sciences 105 (26): 8989–8993, doi:, PMID 18577593, http://www.pnas.org/cgi/reprint/0800388105v1.pdf
- ^ Fernald, Russell D. (September 2006), "Casting a Genetic Light on the Evolution of Eyes", Science 313 (5795): 1914–1918, doi:, PMID 17008522
- ^ a b c Wilson, M. (1978), "The functional organisation of locust ocelli", Journal of Comparative Physiology (4): 297–316
- ^ Wagner, H.J., Douglas, R.H., Frank, T.M., Roberts, N.W., and Partridge, J.C. (Jan. 27, 2009). "A Novel Vertebrate Eye Using Both Refractive and Reflective Optics". Current Biology 19: 108–114. doi:. PMID 19110427.
- ^ Völkel, R.; Eisner, M.; Weible, K. J. (June 2003). "Miniaturized imaging systems" (PDF). Microelectronic Engineering 67-68 (1): 461–472. doi:. http://www.suss-microoptics.com/downloads/Publications/Miniaturized_Imaging_Systems.pdf.
- ^ Gaten, Edward (1998). "Optics and phylogeny: is there an insight? The evolution of superposition eyes in the Decapoda (Crustacea)". Contributions to Zoology 67 (4): 223–236. http://dpc.uba.uva.nl/ctz/vol67/nr04/art01#FIGURE1.
- ^ Ritchie, Alexander (1985). "Ainiktozoon loganense Scourfield, a protochordate? from the Silurian of Scotland". Alcheringa 9: 137.
- ^ Mayer, G. (2006), "Structure and development of onychophoran eyes: What is the ancestral visual organ in arthropods?", Arthropod Structure and Development 35 (4): 231–245, doi:
- ^ a b c d e f Land, M. F. (1989), "The eyes of hyperiid amphipods: relations of optical structure to depth" (PDF), Journal of Comparative Physiology A: Sensory, Neural, and Behavioral Physiology 164 (6): 751–762, doi:, http://www.springerlink.com/index/P0P467K474307K3N.pdf
- ^ Zeil, J. (1996), "The variation of resolution and of ommatidial dimensions in the compound eyes of the fiddler crab Uca lactea annulipes (Ocypodidae, Brachyura, Decapoda)" (PDF), Journal of Experimental Biology 199 (7): 1569–1577, http://jeb.biologists.org/cgi/reprint/199/7/1569.pdf
- ^ John C. Russ (2006). The Image Processing Handbook. CRC Press. ISBN 0849372542. OCLC 156223054. http://books.google.com/books?id=Vs2AM2cWl1AC&pg=PT110&dq=%2250+cycles+per+degree%22+acuity&as_brr=3&ei=i0TESKL7IofMtAPQyYjYDA&sig=ACfU3U31mhrHfaL4ynsg5LzXvGWm7JORiA. "The upper limit (finest detail) visible with the human eye is about 50 cycles per degree,… (Fifth Edition, 2007, Page 94)"
- ^ Curtis D. Klaassen (2001). Casarett and Doull's Toxicology: The Basic Science of Poisons. McGraw-Hill Professional. ISBN 0071347216. OCLC 47965382. http://books.google.com/books?id=G16riRjvmykC&pg=PA574&dq=cycles-per-degree+acuity+rat&as_brr=3&ei=i_8zR_iPKZu8swPhpty3AQ&sig=sI3lzlLANn2lgXbnmRfHH66b6Vw.
- ^ Robert E. Fischer; Biljana Tadic-Galeb. With contributions by Rick Plympton… (2000). Optical System Design. McGraw-Hill Professional. ISBN 0071349162. OCLC 247851267. http://books.google.com/books?id=byx2Ne9cD1IC&pg=PA164&ots=bPLQRko6xA&dq=eye+resolution+line-pairs+1.7&sig=eA2LwmFzvZ9mv-nzZaEINXiWKJE#PPA164,M1.
- ^ Barlow, Horace Basil; Mollon, J. D. (1982), The Senses, pp. 98, ISBN 0521244749, http://books.google.com/books?id=kno6AAAAIAAJ&pg=PA98&vq=human+spectral+sensitivity&dq=eye+visible+spectrum&lr=&as_brr=3&source=gbs_search_s&cad=0
- ^ Fernald, Russell D. (1997), "The Evolution of Eyes" (Free full text), Brain, Behavior and Evolution 50 (4): 253–259, doi:, http://www.stanford.edu/group/fernaldlab/pubs/1997%20Fernald.pdf
- ^ a b c d e Goldsmith, T. H. (1990), "Optimization, Constraint, and History in the Evolution of Eyes" (PDF), The Quarterly Review of Biology 65 (3): 281, doi:, http://www.jstor.org/stable/pdfplus/2832368.pdf
- ^ a b c d e f Nilsson, D (2008). "Eye Evolution: the Blurry Beginning". Current Biology 18: R1096. doi:.
- "Anatomy". History of Ophthalmology. http://www.mrcophth.com/Historyofophthalmology/anatomy.htm. Retrieved on 23 April 2005.
- Kandel ER, Schwartz JH, Jessell TM. Principles of Neural Science, 4th ed. McGraw-Hill, New York (2000). ISBN 0-8385-7701-6
- ^ The precise number depends on the author
[edit] External links
| Wikimedia Commons has media related to: Eyes |
- A human eye
- Evolution of the eye
- Diagram of the eye
- Webvision. The organisation of the retina and visual system. An in-depth treatment of retinal function, open to all but geared most toward graduate students.
- Eye strips images of all but bare essentials before sending visual information to brain, UC Berkeley research shows


{kind=link}