Semantic memory
From Wikipedia, the free encyclopedia
Semantic memory refers to the memory of meanings, understandings, and other concept-based knowledge unrelated to specific experiences. The conscious recollection of factual information and general knowledge about the world,[1] generally thought to be independent of context and personal relevance. Semantic and episodic memory together make up the category of declarative memory, which is one of the two major divisions in memory. The counterpart to declarative, or explicit memory, is procedural memory, or implicit memory.[2]
Semantic memory includes generalized knowledge that does not involve memory of a specific event. For instance, you can answer a question like "Are wrenches pets or tools?" without remembering any specific event in which you learned that wrenches are tools.
Contents |
[edit] History
The notion of semantic memory was first introduced following a conference in 1972 between Endel Tulving, of the University of Toronto, and W. Donaldson on the role of organization in human memory. Tulving constructed a proposal to distinguish between episodic memory and what he termed semantic memory. He was mainly influenced by the ideas of Reiff and Scheers, who in 1959 made the distinction between two primary forms of memory. One form titled remembrances and the other memoria. The remembrance concept dealt with memories that contained the experiences of an autobiographic index, whereas the memoria’ concept dealt with those memories without the experiences of an autobiographic index. Semantic memory was to reflect our knowledge of the world around us. It holds generic information that is more than likely acquired across various contexts and is able to be used across different situations. According to Madigan in his book titled Memory, semantic memory is the sum of all knowledge you have obtained- whether it be your vocabulary, understanding of math, and all the facts you know. The use of semantic memory is quite different from that of episodic memory. Semantic memory refers to general facts and meanings we share with others whereas episodic memory refers to unique and concrete personal experiences. Tulving's proposal of this distinction between semantic and episodic memory was widely accepted mainly because it allowed the separate conceptualization of knowledge of the world. Tulving discusses these separate systems of conceptualization of episodic and semantic memory in his book titled Elements of Episodic Memory. He states that both episodic and semantic memory differ in regards to several factors including:
- the characteristics of their operations,
- the kind of information they process, and
- their application to the real world as well as the memory laboratory.
Before this proposal by Tulving this area of human memory had been neglected by experimental psychologists. A number of experimenters have conducted tests to determine the validity of Tulving’s hypothesized distinction of episodic and semantic memory.
[edit] Empirical evidence
[edit] Kihlstrom (1980)- Experiment 1
In this study four groups of University students, varying in their levels of hypnotic susceptibility, were hypnotized. While under hypnosis they learned a list of 16 common words using a multi-trial free recall method. Once the subjects were able to perfectly recall the list twice in succession they were told that after awakening they would not remember having learned any of the words while under hypnosis. However, given the signal of the experimenter not only will they remember having learned the words but they will also remember the words from the list.
During stage one of the experiment (after subjects were awakened) the number of words recalled by the subjects were used as a measure of performance for the episodic task of free recall. Most subjects remembered learning the list of words.
During the second stage the measure of semantic memory performance was assessed. Each subject was given a semantic free association test (where stimulus words were given to elicit the learned words).
As mentioned previously, the subjects represented various levels of hypnotic susceptibility as determined by their scores on the Stanford Hypnotic Susceptibility Scale. They were grouped according to their score.
The semantic free association probabilities were relatively the same across various hypnotized groups. However, the episodic free recall probabilities were significantly different across the groups. The percentage increased as the hypnotizability of subjects decreased. The subjects in the very high susceptibility group recalled almost nothing, whereas the medium and low groups recalled 86% of the learned words.
Because the free association test was not related to the hypnotic susceptibility of the subjects shows that amnesia presented after hypnosis determined the memory for the word-events that occurred in the study phase.
This study provides evidence that supports the episodic/semantic distinction hypothesized by Tulving.
[edit] Jacob and Dallas (1981)
This study was not created to solely provide evidence for the distinction of semantic and episodic memory stores. However, they did use the experimental dissociation method which provides evidence for Tulving’s hypothesis.
Part one
Subjects were presented with 60 words (one at a time) and were asked different questions.
- Some questions asked were to cause the subject to pay attention to the visual appearance: Is the word typed in bold letters?
- Some questions caused the participants to pay attention to the sound of the word: Does the word rhyme with ball?
- Some questions caused the subjects to pay attention to the meaning of the word: Does the word refer to a form of communication?
- Half of the questions were “no” answers and the other half “yes”
Part Two
In the second phase of the experiment 60 “old words”- seen in stage one and “20 new words” not shown in stage one were presented to the subjects one at a time.
The subjects were given one of two tasks:
- Perceptual Identification task (semantic): The words were flashed on a video-screen for 35ms and the subjects were required to say what the word was.
- Episodic Recognition Task: Subjects were presented with each word and had to decide whether they had seen the word in the previous stage of the experiment.
Results:
- The percentages correct in the Semantic task (perceptual identification) did not change with the encoding conditions of appearance, sound, or meaning.
- The percentages for the episodic task increased from the appearance condition (.50), to the sound condition (.63), to the meaning condition (.86). - The effect was also greater for the “yes” encoding words than the “no” encoding words. (see stage one)
Conclusion: It displays a strong distinction of performance of episodic and semantic tasks, thus supporting Tulving’s hypothesis.
[edit] Models
The essence of semantic memory is that its contents are not tied to any particular instance of experience, as in episodic memory. Instead, what is stored in semantic memory is the "gist" of experience, an abstract structure that applies to a wide variety of experiential objects and which may be said to delineate categorical and functional relationships between such objects.[3] Thus, a complete theory of semantic memory must account not only for the representational structure of such "gists", but also for how they can be extracted from experience. Numerous models of semantic memory have been proposed; they are summarized below.
[edit] Network models
Networks of various sorts play an integral part in many theories of semantic memory. Generally speaking, a network is composed of a set of nodes connected by links. The nodes may represent concepts, words, perceptual features, or nothing at all. The links may be weighted such that some are stronger than others or, equivalently, have a length such that some links take longer to traverse than others. All these features of networks have been employed in models of semantic memory, examples of which are found below.
[edit] Teachable Language Comprehender (TLC)
One of the first examples of a network model of semantic memory is the Teachable Language Comprehender (TLC).[4] In this model, each node is a word, representing a concept (like "Bird"). With each node is stored a set of properties (like "can fly" or "has wings") as well as pointers (i.e., links) to other nodes (like "Chicken"). A node is directly linked to those nodes of which it is either a subclass or superclass (i.e., "Bird" would be connected to both "Chicken" and "Animal"). Thus, TLC is a hierarchical knowledge representation in that high-level nodes representing large categories are connected (directly or indirectly, via the nodes of subclasses) to many instances of those categories, whereas nodes representing specific instances are at a lower level, connected only to their superclasses. Furthermore, properties are stored at the highest category level to which they apply. For example, "is yellow" would be stored with "Canary", "has wings" would be stored with "Bird" (one level up), and "can move" would be stored with "Animal" (another level up). Nodes may also store negations of the properties of their superordinate nodes (i.e., "NOT-can fly" would be stored with "penguin"). This provides an economy of representation in that properties are only stored at the category level at which they become essential, that is, at which point they become critical features (see below).
Processing in TLC is a form of spreading activation.[5] That is, when a node becomes active, that activation spreads to other nodes via the links between them. In that case, the time to answer the question "Is a chicken a bird?" is a function of how far the activation between the nodes for "Chicken" and "Bird" must spread, i.e., the number of links between the nodes "Chicken" and "Bird".
The original version of TLC did not put weights on the links between nodes. This version performed comparably to humans in many tasks, but failed to predict that people would respond faster to questions regarding more typical category instances than those involving less typical instances.[6] Collins and Quillian later updated TLC to include weighted connections to account for this effect.[7] This updated TLC is capable of explaining both the familiarity effect and the typicality effect. Its biggest advantage is that it clearly explains priming: you are more likely to retrieve information from memory if related information (the "prime") has been presented a short time before. There are still a number of memory phenomena for which TLC has no account, including why people are able to respond quickly to obviously false questions (like "is a chicken a meteor?"), when the relevant nodes are very far apart in the network.[8]
[edit] Semantic networks
TLC is an instance of a more general class of models known as semantic networks. In a semantic network, each node is to be interpreted as representing a specific concept, word, or feature. That is, each node is a symbol. Semantic networks generally do not employ distributed representations for concepts, as may be found in a neural network. The defining feature of a semantic network is that its links are almost always directed (that is, they only point in one direction, from a base to a target) and the links come in many different types, each one standing for a particular relationship that can hold between any two nodes.[9] Processing in a semantic network often takes the form of spreading activation (see above).
Semantic networks see the most use in models of discourse and logical comprehension, as well as in Artificial Intelligence.[10] In these models, the nodes correspond to words or word stems and the links represent syntactic relations between them. For an example of a computational implementation of semantic networks in knowledge representation, see Cravo and Martins (1993).[11]
[edit] Feature models
One may view semantic categories as being composed of relatively unstructured sets of features. The semantic feature-comparison model, proposed by Smith, Shoben, and Rips (1974),[12] describes memory as being composed of feature lists for different concepts. According to this view, the relations between categories would not be directly retrieved, they would be indirectly computed. For example, subjects might verify a sentence by comparing the feature sets that represent its subject and predicate concepts. Such computational feature-comparison models include the ones proposed by Meyer (1970),[13] Rips (1975),[14] Smith, et al. (1974).[15]
Early work in perceptual and conceptual categorization assumed that categories had critical features and that category membership could be determined by logical rules for the combination of features. More recent theories have accepted that categories may have an ill-defined or "fuzzy" structure[16] and have proposed probabilistic or global similarity models for the verification of category membership.[17]
[edit] Associative models
The "association"—a relationship between two pieces of information—is a fundamental concept in psychology, and associations at various levels of mental representation are essential to models of memory and cognition in general. The set of associations among a collection of items in memory is equivalent to the links between nodes in a network, where each node corresponds to a unique item in memory. Indeed, neural networks and semantic networks may be characterized as associative models of cognition. However, associations are often more clearly represented as an N×N matrix, where N is the number of items in memory. Thus, each cell of the matrix corresponds to the strength of the association between the row item and the column item.
Learning of associations is generally believed to be a Hebbian process; that is, whenever two items in memory are simultaneously active, the association between them grows stronger, and the more likely either item is to activate the other. See below for specific operationalizations of associative models.
[edit] Search of Associative Memory (SAM)
A standard model of memory that employs association in this manner is the Search of Associative Memory (SAM) model.[18] Though SAM was originally designed to model episodic memory, its mechanisms are sufficient to support some semantic memory representations, as well.[19] In SAM, when any two items simultaneously occupy a working memory buffer, the strength of their association is incremented. Thus, items that co-occur more often are more strongly associated. Items in SAM are also associated with a specific context, where the strength of that association determined by how long each item is present in a given context. In SAM, then, memories consist of a set of associations between items in memory and between items and contexts. The presence of a set of items and/or a context is more likely to evoke, then, some subset of the items in memory. The degree to which items evoke one another—either by virtue of their shared context or their co-occurrence—is an indication of the items’ semantic relatedness.
[edit] ACT-R: a production system model
The ACT (Adaptive Control of Thought)[20] (and later ACT-R (Adaptive Control of Thought-Rational)[21]) theory of cognition represents declarative memory (of which semantic memory is a part) with "chunks", which consist of a label, a set of defined relationships to other chunks (i.e., "this is a _", or "this has a _"), and any number of chunk-specific properties. Chunks, then, can be mapped as a semantic network, given that each node is a chunk with its unique properties, and each link is the chunk’s relationship to another chunk. In ACT, a chunk’s activation decreases as a function of the time since the chunk was created and increases with the number of times the chunk has been retrieved from memory. Chunks can also receive activation from Gaussian noise, and from their similarity to other chunks. For example, if "chicken" is used as a retrieval cue, "canary" will receive activation by virtue of its similarity to the cue (i.e., both are birds, etc.). When retrieving items from memory, ACT looks at the most active chunk in memory; if it is above threshold, it is retrieved, otherwise an "error of omission" has occurred, i.e., the item has been forgotten. There is, additionally, a retrieval latency, which varies inversely with the amount by which the activation of the retrieved chunk exceeds the retrieval threshold. This latency is used in measuring the response time of the ACT model, to compare it to human performance.[22]
While ACT is a model of cognition in general, and not memory in particular, it nonetheless posits certain features of the structure of memory, as described above. In particular, ACT models memory as a set of related symbolic chunks which may be accessed by retrieval cues. While the model of memory employed in ACT is similar in some ways to a semantic network, the processing involved is more akin to an associative model.
[edit] Statistical models
Some models characterize the acquisition of semantic information as a form of statistical inference from a set of discrete experiences, distributed across a number of "contexts". Though these models differ in specifics, they generally employ an (Item × Context) matrix where each cell represents the number of times an item in memory has occurred in a given context. Semantic information is gleaned by performing a statistical analysis of this matrix.
Many of these models bear similarity to the algorithms used in search engines (for example, see Griffiths, et al., 2007[23] and Anderson, 1990[24]), though it is not yet clear whether they really use the same computational mechanisms.
[edit] Latent Semantic Analysis (LSA)
Perhaps the most popular of these models is Latent Semantic Analysis (LSA).[25] In LSA, a T × D matrix is constructed from a text corpus where T is the number of terms in the corpus and D is the number of documents (here "context" is interpreted as "document" and only words—or word phrases—are considered as items in memory). Each cell in the matrix is then transformed according to the equation:
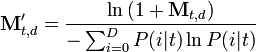
where P(i | t) is the probability that context i is active, given that item t has occurred (this is obtained simply by dividing the raw frequency, 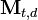 by the total of the item vector,
by the total of the item vector, 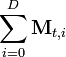 ). This transformation—applying the logarithm, then dividing by the entropy of the item over all contexts—provides for greater differentiation between items and effectively weights items by their ability to predict context, and vice versa (that is, items that appear across many contexts, like "the" or "and", will be weighted less, reflecting their lack of semantic information). A Singular Value Decomposition (SVD) is then performed on the matrix
). This transformation—applying the logarithm, then dividing by the entropy of the item over all contexts—provides for greater differentiation between items and effectively weights items by their ability to predict context, and vice versa (that is, items that appear across many contexts, like "the" or "and", will be weighted less, reflecting their lack of semantic information). A Singular Value Decomposition (SVD) is then performed on the matrix  , which allows the number of dimensions in the matrix to be reduced, thus clustering LSA's semantic representations and providing for indirect association between items. For example, "cat" and "dog" may never appear together in the same context, so their close semantic relationship may not be well-captured by LSA's original matrix
, which allows the number of dimensions in the matrix to be reduced, thus clustering LSA's semantic representations and providing for indirect association between items. For example, "cat" and "dog" may never appear together in the same context, so their close semantic relationship may not be well-captured by LSA's original matrix  . However, by performing the SVD and reducing the number of dimensions in the matrix, the context vectors of "cat" and "dog"—which would be very similar—would migrate toward one another and perhaps merge, thus allowing "cat" and "dog" to act as retrieval cues for each other, even though they may never have co-occurred. The degree of semantic relatedness of items in memory is given by the cosine of the angle between the items' context vectors (ranging from 1 for perfect synonyms to 0 for no relationship). Essentially, then, two words are closely semantically related if they appear in similar types of documents.
. However, by performing the SVD and reducing the number of dimensions in the matrix, the context vectors of "cat" and "dog"—which would be very similar—would migrate toward one another and perhaps merge, thus allowing "cat" and "dog" to act as retrieval cues for each other, even though they may never have co-occurred. The degree of semantic relatedness of items in memory is given by the cosine of the angle between the items' context vectors (ranging from 1 for perfect synonyms to 0 for no relationship). Essentially, then, two words are closely semantically related if they appear in similar types of documents.
[edit] Hyperspace Analogue to Language (HAL)
The Hyperspace Analogue to Language (HAL) model[26][27] considers context only as the words that immediately surround a given word. HAL computes an NxN matrix, where N is the number of words in its lexicon, using a 10-word reading frame that moves incrementally through a corpus of text. Like in SAM (see above), any time two words are simultaneously in the frame, the association between them is increased, that is, the corresponding cell in the NxN matrix is incremented. The amount by which the association is incremented varies inversely with the distance between the two words in the frame (specifically, Δ = 11 − d, where d is the distance between the two words in the frame). As in LSA (see above), the semantic similarity between two words is given by the cosine of the angle between their vectors (dimension reduction may be performed on this matrix, as well). In HAL, then, two words are semantically related if they tend to appear with the same words. Note that this may hold true even when the words being compared never actually co-occur (i.e., "chicken" and "canary").
[edit] Location of semantic memory in the brain
The cognitive neuroscience of semantic memory is a somewhat controversial issue with two dominant views.
On the one hand, many researchers and clinicians believe that semantic memory is stored by the same brain systems involved in episodic memory. These include the medial temporal lobes (MTL) and hippocampal formation. In this system, the hippocampal formation "encodes" memories, or makes it possible for memories to form at all, and the cortex stores memories after the initial encoding process is completed.
Recently, new evidence has been presented in support of a more precise interpretation of this hypothesis. The hippocampal formation includes, among other structures: the hippocampus itself, the entorhinal cortex, and the perirhinal cortex. These latter two make up the "parahippocampal cortices". Amnesics with damage to the hippocampus but some spared parahippocampal cortex were able to demonstrate some degree of intact semantic memory despite a total loss of episodic memory. This strongly suggests that encoding of information leading to semantic memory does not have its physiological basis in the hippocampus.[28]
Other researchers believe the hippocampus is only involved in episodic memory and spatial cognition. This then raises the question where semantic memory may be located. Some believe semantic memory lives in temporal neocortex. Others believe that semantic knowledge is widely distributed across all brain areas. To illustrate this latter view, consider your knowledge of dogs. Researchers holding the 'distributed semantic knowledge' view believe that your knowledge of the sound a dog makes exists in your auditory cortex, whilst your ability to recognize and imagine the visual features of a dog resides in your visual cortex. Perhaps all these representations are indexed by the left temporal pole, a region particularly vulnerable to damage in semantic dementia.
[edit] Neural correlates and biological workings
The hippocampal areas are important to semantic memory's involvement with declarative memory. The left inferior prefrontal cortex (PFC) and the left posterior temporal areas are other areas involved in semantic memory use. Temporal lobe damage affecting the lateral and medial cortexes have been related to semantic impairments. Damage to different areas of the brain affect semantic memory differently.[29]
A brain scan indicates that the left hippocampal areas show an increase in activity during use. During semantic retrieval, two regions in the right middle frontal gyrus and the area of the right inferior temporal gyrus similarly show an increase in activity. Damage to areas involved in semantic memory result in different effects, depending on the area and type of damage. For instance, Lambon Ralph, Lowe, & Rogers (2007) found that category specific impairments can occur where patients have different knowledge deficits for one semantic category over another, depending on location and type of damage. Category-specific impairments might indicate that knowledge may rely differentially upon sensory and motor properties encoded in separate areas (Farah and McClelland, 1991).[30]
Category-specific impairments can involve cortical regions where living and nonliving things are represented and where feature and conceptual relationships are represented. Depending on the damage to the semantic system, one type might be favored over the other. In many cases, there is a point where one domain is better than the other (ie - representation of living and nonliving things over feature and conceptual relationships or vice versa)[31]
Different diseases and disorders can affect the biological workings of semantic memory. A variety of studies have been done in an attempt to determine the effects on varying aspects of semantic memory. For example, Lambon, Lowe, & Rogers (2007) studied the different effects semantic dementia and herpes simplex virus encephalitis have on semantic memory. They found that semantic dementia has a more generalized semantic impairment. Additionally, deficits in semantic memory as a result of herpes simplex virus encephalitis tend to have more category-specific impairments. Other disorders that affect semantic memory - such as Alzheimer's disease - has been observed clinically as errors in naming, recognizing, or describing objects. Whereas researchers have attributed such impairment to degradation of semantic knowledge (Koenig et al 2007).[32]
Various neural imaging and research points to semantic memory and episodic memory resulting from distinct areas in the brain. Still other research suggests that both semantic memory and episodic memory are part of a singular declarative memory system, yet represent different sectors and parts within the greater whole. Different areas within the brain are activated depending on whether semantic or episodic memory is accessed. Certain experts are still arguing whether or not the two types of memory are from distinct systems or whether the neural imaging makes it appear that way as a result of the activation of different mental processes during retrieval.[33]
[edit] Disorders
In order to understand semantic memory disorders, one must first understand how these disorders affect memory. Semantic memory disorders fractionate into two categories. Semantic category specific impairments and modality specific impairments are apparent in disorders of semantic memory. Understanding these types of impairments will give insight into how disorders of semantic memory function.
[edit] Semantic category specific impairments
Category specific impairments can result in widespread, patchy damage or localized damage. Category specific impairments can be broken down into four categories. Perceptual and functional features, topographic organization, informativeness and intercorrelations are areas of decreased functioning in disorders of semantic memory (Warrington and Shallice, 1984).[34] Alzheimer's disease is a semantic memory disorder characterized by category specific impairments. Alzheimer's disease results in naming errors as well as errors describing and naming objects, though not necessarily category-specific[35].Semantic dementia is another disorder associated with semantic memory. Semantic dementia is a language disorder characterized by a deterioration in understanding and recognizing words. Impairments include difficulty in generating familiar words, difficulty naming objects and difficulties with visual recognition. Research suggests that the temporal lobe might be responsible for category specific impairments of semantic memory disorders. In addition to category specific impairments, modality specific impairments are included in disorders of semantic memory (Cohen et al. 2002).[36]
[edit] Modality specific impairments
Semantic memory is also discussed in reference to modality. Different components represent information from different sensorimotor channels. Modality specific impairments are divided into separate subsystems on the basis of input modality. Examples of different input modalities include visual, auditory and tactile input. Modality specific impairments are also divided into subsystems based on the type of information. Visual vs. verbal and perceptual vs. functional information are examples of information types.[37] Modality specificity can account for category specific impairments in semantic memory disorders. Damage to visual semantics primarily impairs knowledge of living things, and damage to functional semantics primarily impairs knowledge of nonliving things.
[edit] Semantic refractory access and semantic storage disorders
Semantic memory disorders fall into two groups. Semantic refractory access disorders are contrasted with semantic storage disorders according to four factors. Temporal factors, response consistency, frequency and semantic relatedness are the four factors used to differentiate between semantic refractory access and semantic storage disorders. A key feature of semantic refractory access disorders is temporal distortions. Decreases in response time to certain stimuli are noted when compared to natural response times. Response consistency is the next factor. In access disorders you see inconsistencies in comprehending and responding to stimuli that have been presented many times. Temporal factors impact response consistency. In storage disorders, you do not see an inconsistent response to specific items like you do in refractory access disorders. Stimulus frequency determines performance at all stages of cognition. Extreme word frequency effects are common in semantic storage disorders while in semantic refractory access disorders word frequency effects are minimal. The comparison of 'close' and 'distant' groups tests semantic relatedness. 'Close' groupings have words that are related because they are drawn from the same category. For example, a listing of clothing types would be a 'close' grouping. 'Distant' groupings contain words with broad categorical differences. Non-related words would fall into this group. Comparing close and distant groups shows that in access disorders semantic relatedness had a negative effect. This is not observed in semantic storage disorders. Category specific and modality specific impairments are important components in access and storage disorders of semantic memory.[38]
[edit] Present and future research
Semantic memory has had a comeback in interest in the past 15 years, due in part to the development of functional neuroimaging methods such as positron emission tomography (PET) and functional magnetic resonance imaging (fMRI), which have been used to address some of the central questions about our understanding of semantic memory.
Rather than any one brain region playing a dedicated and privileged role in the representation or retrieval of all sorts of semantic knowledge, semantic memory is a collection of functionally and anatomically distinct systems, where each attribute-specific system is tied to a sensorimotor modality (i.e. vision) and even more specifically to a property within that modality (i.e. color). Neuroimaging studies also suggest a distinction between semantic processing and sensorimotor processing.
A new idea that is still at the early stages of development is that semantic memory, like perception, can be subdivided into types of visual information – color, size, form, and motion. Thompson-Schill (2003)[39] found that the left or bilateral ventral temporal cortex appears to be involved in retrieval of knowledge of color and form, the left lateral temporal cortex in knowledge of motion, and the parietal cortex in knowledge of size.
Neuroimaging studies suggest a large, distributed network of semantic representations that are organized minimally by attribute, and perhaps additionally by category. These networks include "extensive regions of ventral (form and color knowledge) and lateral (motion knowledge) temporal cortex, parietal cortex (size knowledge), and premotor cortex (manipulation knowledge). Other areas, such as more anterior regions of temporal cortex, may be involved in the representation of nonperceptual (e.g. verbal) conceptual knowledge, perhaps in some categorically-organized fashion."[40]
[edit] References
- ^ "THE BRAIN FROM TOP TO BOTTOM". http://thebrain.mcgill.ca/flash/a/a_07/a_07_p/a_07_p_tra/a_07_p_tra.html#3.
- ^ Tulving, E., & Schacter, D.L. (1990). Priming and human memory systems. Science, 247, 301 – 306.
- ^ Kintsch, W. (1988). The role of knowledge in discourse comprehension: A construction-integration model. Psychological Review, 95, 163–182.
- ^ Collins, A. M. & Quillian, M. R. (1969). Retrieval time from semantic memory. Journal of Verbal Learning and Verbal Behavior, 8, 240-247
- ^ Collins, A. M. & Quillian, M. R. (1972). How to make a language user. In E. Tulving & W. Donaldson (Eds.), Organization of memory (pp. 309-351). New York: Academic Press.
- ^ Rips, L. J., Shoben, E. J. & Smith, F. E. (1973). Semantic distance and the verification of semantic relations. Journal of Verbal Learning and Verbal Behavior, 14, 665-681.
- ^ Collins, A. M. & Loftus, E. F. (1975). A spreading-activation theory of semantic processing. Psychological Review, 82(6), 407-428
- ^ Glass, A. L., Holyoak, K. J. & Kiger, J. I. (1979). Role of antonymy relations in semantic judgments. Journal of Experimental Psychology: Human Learning & Memory, 5(6), 598-606.
- ^ Arbib, M. A. (Ed.). (2002). Semantic networks. In The Handbook of Brain Theory and Neural Networks (2nd ed.), Cambridge, MA: MIT Press.
- ^ Barr, A. & Feigenbaum, E. A. (1982). The handbook of artificial intelligence. Lost Altos, CA: William Kaufman.
- ^ Cravo, M. R. & Martins, J. P. (1993). SNePSwD: A newcomer to the SNePS family. Journal of Experimental & Theoretical Artificial Intelligence, 5, 135–148.
- ^ Smith, E. E., Shoben, E. J. & Rips, L. J. (1974). Structure and process in semantic memory: A featural model for semantic decisions. Psychological Review, 1, 214-241.
- ^ Meyer, D. E. (1970). On the representation and retrieval of stored semantic information. Cognitive Psychology, 1(3), 242-299
- ^ Rips, L. J. (1975). Inductive judgments about natural categories. Journal of Verbal Learning & Verbal Behavior, 14(6), 665-681.
- ^ Smith, E. E., Shoben, E. J. & Rips, L. J. (1974). Structure and process in semantic memory: A featural model for semantic decisions. Psychological Review, 1, 214-241.
- ^ McCloskey, M. E. & Glucksberg, S. (1978). Natural categories: Well defined or fuzzy sets? Memory & Cognition, 6(4), 462-472.
- ^ McCloskey, M. & Glucksberg, S. (1979). Decision processes in verifying category membership statements: Implications for models of semantic memory. Cognitive Psychology, 11(1), 1-37.
- ^ Raaijmakers, J. G. W.; Schiffrin, R. M. (1981), "Search of associative memory", Psychological Review 8 (2): 98-134
- ^ Kimball, D. R., Smith, T. A. & Kahana, M. J. (in press). The fSAM model of false recall. Psychological Review.
- ^ Anderson, J. R. (1983). The Architecture of Cognition. Cambridge, MA: Harvard University Press.
- ^ Anderson, J. R. (1993b). Rules of the mind. Hillsdale, NJ: Erlbaum.
- ^ Anderson, J. R., Bothell, D., Lebiere, C. & Matessa, M. (1998). An integrated theory of list memory. Journal of Memory and Language, 38, 341-380.
- ^ Griffiths, T. L., Steyvers, M. & Firl, A. (2007). Google and the mind: Predicting fluency with PageRank. Psychological Science, 18(12), 1069-1076.
- ^ Anderson, J. R. (1990). The adaptive character of thought. Hillsdale, NJ: Lawrence Erlbaum Associates.
- ^ Landauer, T. K., & Dumais, S. T. (1997). A solution to Plato's problem: The Latent Semantic Analysis theory of the acquisition, induction, and representation of knowledge. Psychological Review, 104, 211-240.
- ^ Lund, K., Burgess, C. & Atchley, R. A. (1995). Semantic and associative priming in a high-dimensional semantic space. Cognitive Science Proceedings (LEA), 660-665.
- ^ Lund, K. & Burgess, C. (1996). Producing high-dimensional semantic spaces from lexical co-occurrence. Behavior Research Methods, Instruments & Computers, 28(2),203-208.
- ^ Vargha-Khadem et al.
- ^ Burianova, H., & Grady, C. L. (2007). Common and Unique Neural Activations in Autobiographical, Episodic, and Semantic Retrieval. Journal of Cognitive Neuroscience, 19(9):1520-34
- ^ Lambon Ralph, M., Lowe, C. & Rogers, T.T. (2007). Neural Basis of Category-specific Semantic Deficits for Living Things: Evidence from semantic dementia, HSVE and a Neural Network Model. Brain: A Journal of Neurology, 130(Pt 4):1127-37.
- ^ Garrard, et. al (2001). Longitudinal Profiles of Semantic Impairment for Living and Nonliving Concepts in Dementia of Alzheimer's Type. Journal of Cognitive Neuroscience, 13(7) 892-909.
- ^ Lambon, R. & Matthew, A. (2007). Neural Basis of Category-specific Semantic Deficits for Living Things: Evidence from semantic dementia, HSVE and a Neural Network Model. Brain: A Journal of Neurology, 130.(Pt 4):1127-37.
- ^ Rajah, M.N. & McIntosh, A.R. (2005). Overlap in the Functional Neural Systems Involved in Semantic and Episodic Memory Retrieval. Journal of Cognitive Neuroscience, 17(3) 470-482.
- ^ Warrington, E. K. & Shallice, T. 1984. Category specific semantic impairments. Brain, 107, 829-853.
- ^ Laws KR, Adlington RL, Gale TM, Moreno-Martinez FJ, Sartori G. A meta-analytic review of category naming in Alzheimer's disease. Neuropsychologia 2007; 45(12):2674-82
- ^ Cohen, G., Johnston, R. & Plunkett, K. (2002). Exploring cognition: Damaged brains and neural networks. Erlbaum: Psychology Press.
- ^ Valentine, T., Brennen, T. & Bredart, S. (1996). The Cognitive psychology of proper names: On the importance of being ernest. London: Routledge.
- ^ McCarthy, R. (1995). Semantic knowledge and semantic representations. Erlbaum: Psychology Press.
- ^ Thompson-Schill, S.L., 2003. Neuroimaging studies of semantic memory: inferring "how" from "where". Neuropsychologia, 41, 280–292.
- ^ Thompson-Schill, S.L., 2003. Neuroimaging studies of semantic memory: inferring "how" from "where". Neuropsychologia, 41, 280–122 .
[edit] External links
- http://www.mtsu.edu/~sschmidt/Cognitive/semantic/semantic.html#VI.%20Conclusions%20on%20Semantic%20Memory
- http://www.newscientist.com/article.ns?id=dn10012&feedId=brain_rss20
- http://diodor.eti.pg.gda.pl An application of computational semantic memory model. Plays 20 questions game on animals domain
|
|||||||||||

