Cell membrane
From Wikipedia, the free encyclopedia
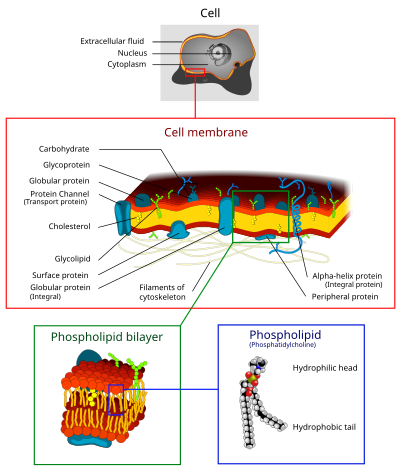
The cell membrane (also called the plasma membrane, plasmalemma, or phospholipid bilayer) is the interface between the cellular machinery inside the cell and the fluid outside.[1]
It is a semipermeable lipid bilayer found in all cells.[2] It contains a wide variety of biological molecules, primarily proteins and lipids, which are involved in a vast array of cellular processes such as cell adhesion, ion channel conductance and cell signaling. The plasma membrane also serves as the attachment point for both the intracellular cytoskeleton and, if present, the extracellular cell wall.
Contents |
Function
The cell membrane surrounds the cytoplasm of a cell and, in animal cells, physically separates the intracellular components from the extracellular environment, thereby serving a function similar to that of skin. In fungi, some bacteria, and plants, an additional cell wall forms the outermost boundary; however, the cell wall plays mostly a mechanical support role rather than a role as a selective boundary. The cell membrane also plays a role in anchoring the cytoskeleton to provide shape to the cell, and in attaching to the extracellular matrix to help group cells together in the formation of tissues. The barrier is selectively permeable and able to regulate what enters and exits the cell, thus facilitating the transport of materials needed for survival. The movement of substances across the membrane can be either passive, occurring without the input of cellular energy, or active, requiring the cell to expend energy in moving it. The membrane also maintains the cell potential.
Specific proteins embedded in the cell membrane can act as molecular signals that allow cells to communicate with each other. Protein receptors are found ubiquitously and function to receive signals from both the environment and other cells. These signals are transduced and passed in a different form into the cell. For example, a hormone binding to a receptor could open an ion channel in the receptor and allow calcium ions to flow into the cell. Other proteins on the surface of the cell membrane serve as "markers" that identify a cell to other cells. The interaction of these markers with their respective receptors forms the basis of cell-cell interaction in the immune system.
Structure
Lipid bilayer

The cell membrane consists primarily of a thin layer of amphipathic phospholipids which spontaneously arrange so that the hydrophobic "tail" regions are shielded from the surrounding polar fluid, causing the more hydrophilic "head" regions to associate with the cytosolic and extracellular faces of the resulting bilayer. This forms a continuous, spherical lipid bilayer.
The arrangement of hydrophilic heads and hydrophobic tails of the lipid bilayer prevent polar solutes (e.g. amino acids, nucleic acids, carbohydrates, proteins, and ions) from diffusing across the membrane, but generally allows for the passive diffusion of hydrophobic molecules. This affords the cell the ability to control the movement of these substances via transmembrane protein complexes such as pores and gates.
Flippases and Scramblases concentrate phosphatidyl serine, which carries a negative charge, on the inner membrane. Along with NANA, this creates an extra barrier to charged moieties moving through the membrane.
Membranes serve diverse functions in eukaryotic and prokaryotic cells. One important role is to regulate the movement of materials into and out of cells. The phospholipid bilayer structure (fluid mosaic model) with specific membrane proteins accounts for the selective permeability of the membrane and passive and active transport mechanisms. In addition, membranes in prokaryotes and in the mitochondria and chloroplasts of eukaryotes facilitate the synthesis of ATP through chemiosmosis.
Membrane polarity
The apical membrane of a polarized cell is the part of the plasma membrane that forms its lumenal surface, distinct from the basolateral membrane. This is particularly evident in epithelial and endothelial cells, but also describes other polarized cells, such as neurons.
The basolateral membrane of a polarized cell is the part of the plasma membrane that forms its basal and lateral surfaces, distinct from the Apical membrane (or luminal) surface. This is particularly evident in epithelial cells, but also describes other polarized cells, such as neurons.
"Basolateral membrane" is a compound phrase referring to the terms basal (base) membrane and lateral (side) membrane, which, especially in epithelial cells, are essentially functionally identical in composition and activity. Proteins (such as ion channels and pumps) are free to move from the basal to the lateral surface of the cell or vice versa in accordance with the fluid mosaic model.
Tight junctions that join epithelial cells near their apical surface prevent the migration of proteins to the apical membrane. The basal and lateral surfaces thus remain roughly equivalent to one another, yet distinct from the apical surface.
Integral membrane proteins
The cell membrane contains many integral membrane proteins, which pepper the entire surface. These structures, which can be visualized by electron microscopy or fluorescence microscopy, can be found on the inside of the membrane, the outside, or membrane spanning. These may include integrins, cadherins, desmosomes, clathrin-coated pits, caveolaes, and different structures involved in cell adhesion.
Membrane skeleton
The cytoskeleton is found underlying the cell membrane in the cytoplasm and provides a scaffolding for membrane proteins to anchor to, as well as forming organelles that extend from the cell. Indeed, cytoskeletal elements interact extensively and intimately with the cell membrane.[3] Anchoring proteins restricts them to a particular cell surface — for example, the apical surface of epithelial cells that line the vertebrate gut — and limits how far they may diffuse within the bilayer. The cytoskeleton is able to form appendage-like organelles, such as cilia, which are microtubule-based extensions covered by the cell membrane, and filopodia, which are actin-based extensions. These extensions are ensheathed in membrane and project from the surface of the cell in order to sense the external environment and/or make contact with the substrate or other cells. The apical surfaces of epithelial cells are dense with actin-based finger-like projections known as microvilli, which increase cell surface area and thereby increase the absorption rate of nutrients. Localized decoupling of the cytoskeleton and cell membrane results in formation of a bleb.
Structure and the Fluid mosaic model
|
|
Please help improve this article or section by expanding it. Further information might be found on the talk page. (June 2008) |
According to the fluid mosaic model of S. J. Singer and Garth Nicolson, the biological membranes can be considered as a two-dimensional liquid where all lipid and protein molecules diffuse more or less freely[4]. This picture may be valid in the space scale of 10 nm. However, the plasma membranes contain different structures or domains that can be classified as (a) protein-protein complexes; (b) lipid rafts, (c) pickets and fences formed by the actin-based cytoskeleton; and (d) large stable structures, such as synapses or desmosomes.
The fluid mosaic model can be seen when the membrane proteins of two cells (e.g., a human cell and a mouse cell) are tagged with different-coloured fluorescent labels. When the two cells are fused, the two colours intermix, indicating that the proteins are free to move in the 2D plane.
Proteins in the cell membranes may be integral or peripheral. Peripheral proteins are present on only one side of the membrane, and integral proteins span the entire membrane. The hydrophobic central layer is anchored in the lipid blayer by hydrophobic bonds, and hydrophillic regions protrude into the extracellular and intracellular fluids. The lipid component of the cell membrane is responsible for the permiability of hydrophobic molecules and small uncharge polar molecules to freely pass through. Large uncharged polar molecules and ions need to be moved across the membrane via transporters (carriers) and channels.
Composition
Cell membranes contain a variety of biological molecules, notably lipids and proteins. Material is incorporated into the membrane, or deleted from it, by a variety of mechanisms:
- Fusion of intracellular vesicles with the membrane (exocytosis) not only excretes the contents of the vesicle but also incorporates the vesicle membrane's components into the cell membrane. The membrane may form blebs around extracellular material that pinch off to become vesicles (endocytosis).
- If a membrane is continuous with a tubular structure made of membrane material, then material from the tube can be drawn into the membrane continuously.
- Although the concentration of membrane components in the aqueous phase is low (stable membrane components have low solubility in water), exchange of molecules with this small reservoir is possible.
In all cases, the mechanical tension in the membrane has an effect on the rate of exchange. In some cells, usually having a smooth shape, the membrane tension and area are interrelated by elastic and dynamical mechanical properties, and the time-dependent interrelation is sometimes called homeostasis, area regulation or tension regulation.
Lipids

The cell membrane consists of three classes of amphipathic lipids: phospholipids, glycolipids, and steroids. The amount of each depends upon the type of cell, but in the majority of cases phospholipids are the most abundant.[5] In RBC studies, 30% of the plasma membrane is lipid.
The fatty chains in phospholipids and glycolipids usually contain an even number of carbon atoms, typically between 16 and 20. The 16- and 18-carbon fatty acids are the most common. Fatty acids may be saturated or unsaturated, with the configuration of the double bonds nearly always cis. The length and the degree of unsaturation of fatty acid chains have a profound effect on membranes fluidity[6] as unsaturated lipids create a kink, preventing the fatty acids from packing together as tightly, thus decreasing the melting point (increasing the fluidity) of the membrane. The ability of some organisms to regulate the fluidity of their cell membranes by altering lipid composition is called homeoviscous adaptation.
The entire membrane is held together via non-covalent interaction of hydrophobic tails, however the structure is quite fluid and not fixed rigidly in place. Phospholipid molecules in the cell membrane are "fluid" in the sense that they are free to diffuse and exhibit rapid lateral diffusion along the layer in which they are present. However, movement of phospholipid molecules between layers is not energetically favourable and does not occur to an appreciable extent. Lipid rafts and caveolae are examples of cholesterol-enriched microdomains in the cell membrane.
In animal cells cholesterol is normally found dispersed in varying degrees throughout cell membranes, in the irregular spaces between the hydrophobic tails of the membrane lipids, where it confers a stiffening and strengthening effect on the membrane.[2]
Carbohydrates
Plasma membranes also contain carbohydrates, predominantly glycoproteins, but with some glycolipids (cerebrosides and gangliosides). For the most part, no glycosylation occurs on membranes within the cell, and only ever occurs on the extracellular surface of the plasma membrane.
The glycocalyx is an important feature in all cells, especially epithelia with microvilli. Recent data suggest the glycocalyx participates in cell adhesion, lymphocyte homing, and many others.
The penultimate sugar is galactose and the terminal sugar is sialic acid, as the sugar backbone is modified in the golgi apparatus. Sialic acid carries a negative charge, providing an external barrier to charged particles.
Proteins
| Type | Description | Examples |
| Integral proteins or transmembrane proteins |
Span the membrane and have a hydrophilic cytosolic domain, which interacts with internal molecules, a hydrophobic membrane-spanning domain that anchors it within the cell membrane, and a hydrophilic extracellular domain that interacts with external molecules. The hydrophobic domain consists of one, multiple, or a combination of α-helices and β sheet protein motifs. | Ion channels, proton pumps, G protein-coupled receptor |
| Lipid anchored proteins | Covalently-bound to single or multiple lipid molecules; hydrophobically insert into the cell membrane and anchor the protein. The protein itself is not in contact with the membrane. | G proteins |
| Peripheral proteins | Attached to integral membrane proteins, or associated with peripheral regions of the lipid bilayer. These proteins tend to have only temporary interactions with biological membranes, and, once reacted the molecule, dissociates to carry on its work in the cytoplasm. | Some enzymes, some hormones |
The cell membrane plays host to a large amount of protein that is responsible for its various activities. The amount of protein differs between species and according to function, however the typical amount in a cell membrane is 50%.[6] These proteins are undoubtedly important to a cell: Approximately a third of the genes in yeast code specifically for them, and this number is even higher in multicellular organisms.[5]
The cell membrane, being exposed to the outside environment, is an important site of cell-cell communication. As such, a large variety of protein receptors and identification proteins, such as antigens, are present on the surface of the membrane. Functions of membrane proteins can also include cell-cell contact, surface recognition, cytoskeleton contact, signaling, enzymatic activity, or transporting substances across the membrane.
Most membrane proteins must be inserted in some way into the membrane. For this to occur, an N-terminus "signal sequence" of amino acids directs proteins to the endoplasmic reticulum, which inserts the proteins into a lipid bilayer. Once inserted, the proteins is then transported to its final destination in vesicles, where the vesicle fuses with the target membrane.
Variation
The cell membrane has slightly different composition in different cell types and has therefore different denominations in different cell types:
- Sarcolemma in myocytes
- Oolemma in oocytes.
Permeability
The permeability of membranes is the ease of molecules to pass through it. Permability depends mainly on the electric charge of the molecule and to a lesser extent the molar mass of the molecule. Electrically-neutral and small molecules pass the membrane easier than charged, large ones.
The inability of charged molecules to pass through the cell membrane results in pH parturition of substances throughout the fluid compartments of the body.
See also
- Cell damage, including damage to the cell membrane
- Ammonium transporter
- AP2 adaptors
- Bacterial cell structure
- Cell adhesion
- Efflux (microbiology)
- Elasticity of cell membranes
- Gram-negative bacteria
- Gram-positive bacteria
References
- ^ Kimball's Biology Pages, Cell Membranes
- ^ a b Alberts B, Johnson A, Lewis J, et al (2002). Molecular Biology of the Cell (4th ed.). ISBN 0-8153-3218-1. http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=mboc4.section.1864.
- ^ Doherty GJ and McMahon HT (2008). "Mediation, Modulation and Consequences of Membrane-Cytoskeleton Interactions". Annual Review of Biophysics 37: 65-95. doi:. PMID 18573073. http://arjournals.annualreviews.org/doi/abs/10.1146/annurev.biophys.37.032807.125912.
- ^ Singer SJ, Nicolson GL (Feb 1972). "The fluid mosaic model of the structure of cell membranes". Science 175 (23): 720–31. PMID 4333397. http://www.sciencemag.org/cgi/pmidlookup?view=long&pmid=4333397.
- ^ a b Lodish H, Berk A, Zipursky LS, et al (2004). Molecular Cell Biology (4th ed.). ISBN 0716731363.
- ^ a b Jesse Gray, Shana Groeschler, Tony Le, Zara Gonzalez (2002). "Membrane Structure" (SWF). Davidson College. http://www.bio.davidson.edu/people/macampbell/111/memb-swf/membranes.swf. Retrieved on 2007-01-11.
External links
| Wikimedia Commons has media related to: Cell membrane |
- Lipids, Membranes and Vesicle Trafficking - The Virtual Library of Biochemistry and Cell Biology
- Cell membrane protein extraction protocol
- Membrane homeostasis, tension regulation, mechanosensitive membrane exchange and membrane traffic
- 3D structures of proteins associated with plasma membrane of eukaryotic cells
|
|||||||||||||||||
|
|||||

