Photosynthesis
From Wikipedia, the free encyclopedia

Photosynthesis[α] is a metabolic pathway that converts carbon dioxide into organic compounds, especially sugars, using the energy from sunlight.[1] Photosynthesis occurs in plants, algae, and many species of Bacteria, but not in Archaea. Photosynthetic organisms are called photoautotrophs, but not all organisms that use light as a source of energy carry out photosynthesis, since photoheterotrophs use organic compounds, rather than carbon dioxide, as a source of carbon.[2] In plants, algae and cyanobacteria photosynthesis uses carbon dioxide and water, releasing oxygen as a waste product. Photosynthesis is crucially important for life on Earth, since as well as it maintaining the normal level of oxygen in the atmosphere, nearly all life either depends on it directly as a source of energy, or indirectly as the ultimate source of the energy in their food.[2][β] The amount of energy trapped by photosynthesis is immense, approximately 100 terawatts :[3] which is about six times larger than the yearly power consumption of human civilization.[4] In all, photosynthetic organisms convert around 100,000,000,000 tonnes of carbon into biomass per year.[5]
Although photosynthesis can occur in different ways in different species, some features are always the same. For example, the process always begins when energy from light is absorbed by proteins called photosynthetic reaction centers that contain chlorophyll. In plants these proteins are held inside organelles called chloroplasts, while in bacteria they are embedded in the plasma membrane. Some of the light energy gathered by chlorophylls is stored in the form of adenosine triphosphate (ATP). The rest of the energy is used to remove electrons from a substance such as water. These electrons are then used in the reactions that turn carbon dioxide into organic compounds. In plants, algae and cyanobacteria these reactions are called the Calvin cycle, but different sets of reactions can be found in bacteria, such as the reverse Krebs cycle in Chlorobium. Many photosynthetic organisms have adaptations that concentrate or store carbon dioxide, this helps reduce a wasteful process called photorespiration that would otherwise consume part of the sugar produced during photosynthesis.
Photosynthesis evolved early in the evolutionary history of life, when all forms of life on Earth were microorganisms. Although the dates are difficult to estimate with any accuracy, the first photosynthetic organisms probably evolved about , and used hydrogen or hydrogen sulfide as sources of electrons, rather than water.[6] Cyanobacteria appeared later, around , and changed the Earth forever when they began to oxygenate the atmosphere, beginning about .[7] This new atmosphere allowed the evolution of complex life such as protists. Eventually, about , one of these protists formed a symbiotic relationship with a cyanobacterium, producing the ancestor of the plants and algae.[8] The chloroplasts in modern plants are the descendants of these ancient symbiotic cyanobacteria.
Contents |
Overview

Photosynthetic organisms are photoautotrophs, which means that they are able to synthesize food directly from carbon dioxide using energy from light. In plants, algae and cyanobacteria, photosynthesis releases oxygen, this is called oxygenic photosynthesis. Although there are some differences between oxygenic photosynthesis in plants, algae and cyanobacteria, the overall process is quite similar in these organisms. However, there are some types of bacteria that carry out anoxygenic photosynthesis, which consumes carbon dioxide but does not release oxygen.
Carbon dioxide is converted into sugars in a process called carbon fixation. Carbon fixation is a redox reaction, so photosynthesis needs to supply both a source of energy to drive this process, and also the electrons needed to convert carbon dioxide into carbohydrate, which is a reduction reaction. In general outline, photosynthesis is the opposite of cellular respiration, where glucose and other compounds are oxidized to produce carbon dioxide, water, and release chemical energy. However, the two processes take place through a different sequence of chemical reactions and in different cellular compartments.
The general equation for photosynthesis is therefore:
- CO2 + 2 H2A + photons → (CH2O)n + H2O + 2A
- carbon dioxide + electron donor + light energy → carbohydrate + oxygen + oxidized electron donor
Since water is most often used as the electron donor in oxygenic photosynthesis, the equation for this process is:
- CO2 + 2 H2O + photons → (CH2O)n + H2O + O2
- carbon dioxide + water + light energy → carbohydrate + oxygen + water
Other processes (e.g. as used by microbial species in Mono Lake, California) substitute other compounds (such as arsenite) for water in the electron-supply role; the microbes use sunlight to reduce arsenite to arsenate:[9] The equation for this reaction is:
- (AsO33-) + CO2 + photons → CO + (AsO43-)[10]
- carbon dioxide + arsenite + light energy → arsenate + carbon monoxide (used to build other compounds in subsequent reactions)
Photosynthesis occurs in two stages. In the first stage, light-dependent reactions or light reactions capture the energy of light and use it to make the energy-storage molecules ATP and NADPH. During the second stage, the light-independent reactions use these products to capture and reduce carbon dioxide.
Photosynthetic membranes and organelles
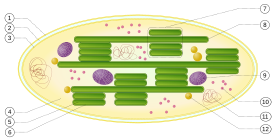
1. outer membrane
2. intermembrane space
3. inner membrane (1+2+3: envelope)
4. stroma (aqueous fluid)
5. thylakoid lumen (inside of thylakoid)
6. thylakoid membrane
7. granum (stack of thylakoids)
8. thylakoid (lamella)
9. starch
10. ribosome
11. plastidial DNA
12. plastoglobule (drop of lipids)
The proteins that gather light for photosynthesis are embedded within cell membranes. The simplest way these are arranged is in photosynthetic bacteria, where these proteins are held within the plasma membrane.[11] However, this membrane may be tightly-folded into cylindrical sheets called thylakoids,[12] or bunched up into round vesicles called intracytoplasmic membranes.[13] These structures can fill most of the interior of a cell, giving the membrane a very large surface area and therefore increasing the amount of light that the bacteria can absorb.[12]
In plants and algae, photosynthesis takes place in organelles called chloroplasts. A chloroplast is contained by an envelope that consists of an inner and an outer phospholipid membrane. Between these two layers is the intermembrane space. A typical plant cell contains about 10 to 100 chloroplasts. Within the stroma are stacks of thylakoids, the sub-organelles which are the site of photosynthesis. The thylakoids are arranged in stacks called grana (singular: granum). A thylakoid has a flattened disk shape. Inside it is an empty area called the thylakoid space or lumen. The thylakoid membrane contains many integral and peripheral membrane proteins. The proteins complexes which contain special pigments absorbing light energy are called photosystems.
Plants absorb light primarily using the pigment chlorophyll, which is the reason that most plants have a green color. Besides chlorophyll plants also use pigments such as carotenes and xanthophylls.[14] Algae also use chlorophyll, but various other pigments are present as phycocyanin, carotenes, and xanthophylls in green algae, phycoerythrin in red algae (rhodophytes) and fucoxanthol in brown algae and diatoms resulting in a wide variety of colors.
These pigments are embedded in plants and algae in special antenna-proteins. In such proteins all the pigments are ordered to work well together. Such a protein is also called a light-harvesting complex.
Although all cells in the green parts of a plant have chloroplasts, most of the energy is captured in the leaves. The cells in the interior tissues of a leaf, called the mesophyll, can contain between 450,000 and 800,000 chloroplasts for every square millimeter of leaf. The surface of the leaf is uniformly coated with a water-resistant waxy cuticle that protects the leaf from excessive evaporation of water and decreases the absorption of ultraviolet or blue light to reduce heating. The transparent epidermis layer allows light to pass through to the palisade mesophyll cells where most of the photosynthesis takes place.
Light reactions

In the light reactions, one molecule of the pigment chlorophyll absorbs one photon and loses one electron. This electron is passed to a modified form of chlorophyll called pheophytin, which passes the electron to a quinone molecule, allowing the start of a flow of electrons down an electron transport chain that leads to the ultimate reduction of NADP to NADPH. In addition, this creates a proton gradient across the chloroplast membrane; its dissipation is used by ATP synthase for the concomitant synthesis of ATP. The chlorophyll molecule regains the lost electron from a water molecule through a process called photolysis, which releases a dioxygen (O2) molecule. The overall equation for the light-dependent reactions under the conditions of non-cyclic electron flow in green plants is:[15]
- 2 H2O + 2 NADP+ + 2 ADP + 2 Pi + light → 2 NADPH + 2 H+ + 2 ATP + O2
Not all wavelengths of light can support photosynthesis. The photosynthetic action spectrum depends on the type of accessory pigments present. For example, in green plants, the action spectrum resembles the absorption spectrum for chlorophylls and carotenoids with peaks for violet-blue and red light. In red algae, the action spectrum overlaps with the absorption spectrum of phycobilins for blue-green light, which allows these algae to grow in deeper waters that filter out the longer wavelengths used by green plants. The non-absorbed part of the light spectrum is what gives photosynthetic organisms their color (e.g., green plants, red algae, purple bacteria) and is the least effective for photosynthesis in the respective organisms.
Z scheme


In plants, light-dependent reactions occur in the thylakoid membranes of the chloroplasts and use light energy to synthesize ATP and NADPH. The light-dependent reaction has two forms; cyclic and non-cyclic reaction. In the non-cyclic reaction, the photons are captured in the light-harvesting antenna complexes of photosystem II by chlorophyll and other accessory pigments (see diagram at right). When a chlorophyll molecule at the core of the photosystem II reaction center obtains sufficient excitation energy from the adjacent antenna pigments, an electron is transferred to the primary electron-acceptor molecule, Pheophytin, through a process called Photoinduced charge separation. These electrons are shuttled through an electron transport chain, the so called Z-scheme shown in the diagram, that initially functions to generate a chemiosmotic potential across the membrane. An ATP synthase enzyme uses the chemiosmotic potential to make ATP during photophosphorylation, whereas NADPH is a product of the terminal redox reaction in the Z-scheme. The electron enters the Photosystem I molecule. The electron is excited due to the light absorbed by the photosystem. A second electron carrier accepts the electron, which again is passed down lowering energies of electron acceptors. The energy created by the electron acceptors is used to move hydrogen ions across the thylakoid membrane into the lumen. The electron is used to reduce the co-enzyme NADP, which has functions in the light-independent reaction. The cyclic reaction is similar to that of the non-cyclic, but differs in the form that it generates only ATP, and no reduced NADP (NADPH) is created. The cyclic reaction takes place only at photosystem I. Once the electron is displaced from the photosystem, the electron is passed down the electron acceptor molecules and returns back to photosystem I, from where it was emitted, hence the name cyclic reaction.
Water photolysis
The NADPH is the main reducing agent in chloroplasts, providing a source of energetic electrons to other reactions. Its production leaves chlorophyll with a deficit of electrons (oxidized), which must be obtained from some other reducing agent. The excited electrons lost from chlorophyll in photosystem I are replaced from the electron transport chain by plastocyanin. However, since photosystem II includes the first steps of the Z-scheme, an external source of electrons is required to reduce its oxidized chlorophyll a molecules. The source of electrons in green-plant and cyanobacterial photosynthesis is water. Two water molecules are oxidized by four successive charge-separation reactions by photosystem II to yield a molecule of diatomic oxygen and four hydrogen ions; the electron yielded in each step is transferred to a redox-active tyrosine residue that then reduces the photoxidized paired-chlorophyll a species called P680 that serves as the primary (light-driven) electron donor in the photosystem II reaction center. The oxidation of water is catalyzed in photosystem II by a redox-active structure that contains four manganese ions and a calcium ion; this oxygen-evolving complex binds two water molecules and stores the four oxidizing equivalents that are required to drive the water-oxidizing reaction. Photosystem II is the only known biological enzyme that carries out this oxidation of water. The hydrogen ions contribute to the transmembrane chemiosmotic potential that leads to ATP synthesis. Oxygen is a waste product of light-dependent reactions, but the majority of organisms on Earth use oxygen for cellular respiration, including photosynthetic organisms.[16][17]
Oxygen and photosynthesis
With respect to oxygen and photosynthesis, there are two important concepts.
- Plant and cyanobacterial (blue-green algae) cells also use oxygen for cellular respiration, although they have a net output of oxygen since much more is produced during photosynthesis.
- Oxygen is a product of the light-driven water-oxidation reaction catalyzed by photosystem II; it is not generated by the fixation of carbon dioxide. Consequently, the source of oxygen during photosynthesis is water, not carbon dioxide.
The concept that oxygen production is not directly associated with the fixation of carbon dioxide was first proposed by Cornelis Van Niel in the 1930s, who studied photosynthetic bacteria. Aside from the cyanobacteria, bacteria only have one photosystem and use reducing agents other than water. They get electrons from a variety of different inorganic chemicals including sulfide or hydrogen, so for most of these bacteria oxygen is not produced.
Others, such as the halophiles (an Archaea), produced so-called purple membranes where the bacteriorhodopsin could harvest light and produce energy. The purple membranes was one of the first to be used to demonstrate the chemiosmotic theory: light hit the membranes and the pH of the solution that contained the purple membranes dropped as protons were pumping out of the membrane.
Light-independent reactions
The Calvin Cycle
In the Light-independent or dark reactions the enzyme RuBisCO captures CO2 from the atmosphere and in a process that requires the newly formed NADPH, called the Calvin-Benson Cycle, releases three-carbon sugars, which are later combined to form sucrose and starch. The overall equation for the light-independent reactions in green plants is:[15]
- 3 CO2 + 9 ATP + 6 NADPH + 6 H+ → C3H6O3-phosphate + 9 ADP + 8 Pi + 6 NADP+ + 3 H2O

To be more specific, carbon fixation produces an intermediate product, which is then converted to the final carbohydrate products. The carbon skeletons produced by photosynthesis are then variously used to form other organic compounds, such as the building material cellulose, as precursors for lipid and amino acid biosynthesis, or as a fuel in cellular respiration. The latter occurs not only in plants but also in animals when the energy from plants gets passed through a food chain.
The fixation or reduction of carbon dioxide is a process in which carbon dioxide combines with a five-carbon sugar, ribulose 1,5-bisphosphate (RuBP), to yield two molecules of a three-carbon compound, glycerate 3-phosphate (GP), also known as 3-phosphoglycerate (PGA). GP, in the presence of ATP and NADPH from the light-dependent stages, is reduced to glyceraldehyde 3-phosphate (G3P). This product is also referred to as 3-phosphoglyceraldehyde (PGAL) or even as triose phosphate. Triose is a 3-carbon sugar (see carbohydrates). Most (5 out of 6 molecules) of the G3P produced is used to regenerate RuBP so the process can continue (see Calvin-Benson cycle). The 1 out of 6 molecules of the triose phosphates not "recycled" often condense to form hexose phosphates, which ultimately yield sucrose, starch and cellulose. The sugars produced during carbon metabolism yield carbon skeletons that can be used for other metabolic reactions like the production of amino acids and lipids.
C4 and C3 photosynthesis and CAM

In hot and dry conditions, plants will close their stomata to prevent loss of water. Under these conditions, CO2 will decrease, and dioxygen gas, produced by the light reactions of photosynthesis, will increase in the leaves, causing an increase of photorespiration by the oxygenase activity of ribulose-1,5-bisphosphate carboxylase/oxygenase and decrease in carbon fixation. Some plants have evolved mechanisms to increase the CO2 concentration in the leaves under these conditions.
C4 plants chemically fix carbon dioxide in the cells of the mesophyll by adding it to the three-carbon molecule phosphoenolpyruvate (PEP), a reaction catalyzed by an enzyme called PEP carboxylase and which creates the four-carbon organic acid, oxaloacetic acid. Oxaloacetic acid or malate synthesized by this process is then translocated to specialized bundle sheath cells where the enzyme, rubisco, and other Calvin cyle enzymes are located, and where CO2 released by decarboxylation of the four-carbon acids is then fixed by rubisco activity to the three-carbon sugar 3-Phosphoglyceric_acids. The physical separation of rubisco from the oxygen-generating light reactions reduces photorespiration and increases CO2 fixation and thus photosynthetic capacity of the leaf. [18] C4 plants can produce more sugar than C3 plants in conditions of high light and temperature. Many important crop plants are C4 plants including maize, sorghum, sugarcane, and millet. Plants lacking PEP-carboxylase are called C3 plants because the primary carboxylation reaction, catalyzed by Rubiso, produces the three-carbon sugar 3-phosphoglyceric acids directly in the Calvin-Benson Cycle.
Xerophytes such as cacti and most succulents also use PEP carboxylase to capture carbon dioxide in a process called Crassulacean acid metabolism (CAM). In contrast to C4 metabolism, which physically separates the CO2 fixation to PEP from the Calvin cycle, CAM only temporally separates these two processes. CAM plants have a different leaf anatomy than C4 plants, and fix the CO2 at night, when their stomata are open. CAM plants store the CO2 mostly in the form of malic acid via carboxylation of phosphoenolpyruvate to oxaloacetate, which is then reduced to malate. Decarboxylation of malate during the day releases CO2 inside the leaves thus allowing carbon fixation to 3-phosphoglycerate by rubisco.
Order and kinetics
The overall process of photosynthesis takes place in four stages. The first, energy transfer in antenna chlorophyll takes place in the femtosecond [1 femtosecond (fs) = 10,−15 s] to picosecond [1 picosecond (ps) = 10−12 s] time scale. The next phase, the transfer of electrons in photochemical reactions, takes place in the picosecond to nanosecond time scale [1 nanosecond (ns) = 10−9 s]. The third phase, the electron transport chain and ATP synthesis, takes place on the microsecond [1 microsecond (μs) = 10−6 s] to millisecond [1 millisecond (ms) = 10−3 s) time scale. The final phase is carbon fixation and export of stable products and takes place in the millisecond to second time scale. The first three stages occur in the thylakoid membranes.
Efficiency
Plants convert light into chemical energy with a maximum photosynthetic efficiency of approximately 6%.[19] By comparison solar panels convert light into electric energy at a photosynthetic efficiency of approximately 10-20%. Actual plant's photosynthetic efficiency varies with the frequency of the light being converted, light intensity, temperature and proportion of CO2 in atmosphere.
Evolution

Early photosynthetic systems, such as those from green and purple sulfur and green and purple non-sulfur bacteria, are thought to have been anoxygenic, using various molecules as electron donors. Green and purple sulfur bacteria are thought to have used hydrogen and sulfur as an electron donor. Green nonsulfur bacteria used various amino and other organic acids. Purple nonsulfur bacteria used a variety of non-specific organic molecules. The use of these molecules is consistent with the geological evidence that the atmosphere was highly reduced at that time.[citation needed]
Fossils of what are thought to be filamentous photosynthetic organisms have been dated at 3.4 billion years old.[20]
The main source of oxygen in the atmosphere is oxygenic photosynthesis, and its first appearance is sometimes referred to as the oxygen catastrophe. Geological evidence suggests that oxygenic photosynthesis, such as that in cyanobacteria, became important during the Paleoproterozoic era around 2 billion years ago. Modern photosynthesis in plants and most photosynthetic prokaryotes is oxygenic. Oxygenic photosynthesis uses water as an electron donor which is oxidized to molecular dioxygen (O2) in the photosynthetic reaction center.
Symbiosis and the origin of chloroplasts
Several groups of animals have formed symbiotic relationships with photosynthetic algae. These are most common in corals, sponges and sea anemones, possibly due to these animals having particularly simple body plans and large surface areas compared to their volumes.[21] In addition, a few marine molluscs Elysia viridis and Elysia chlorotica also maintain a symbiotic relationship with chloroplasts that they capture from the algae in their diet and then store in their bodies. This allows the molluscs to survive solely by photosynthesis for several months at a time.[22][23] Some of the genes from the plant cell nucleus have even been transferred to the slugs, so that the chloroplasts can be supplied with proteins that they need to survive.[24]
An even closer form of symbiosis may explain the origin of chloroplasts. Chloroplasts have many similarities with photosynthetic bacteria including a circular chromosome, prokaryotic-type ribosomes, and similar proteins in the photosynthetic reaction center.[25][26] The endosymbiotic theory suggests that photosynthetic bacteria were acquired (by endocytosis) by early eukaryotic cells to form the first plant cells. Therefore, chloroplasts may be photosynthetic bacteria that adapted to life inside plant cells. Like mitochondria, chloroplasts still possess their own DNA, separate from the nuclear DNA of their plant host cells and the genes in this chloroplast DNA resemble those in cyanobacteria.[27] DNA in chloroplasts codes for redox proteins such as photosynthetic reaction centers. The CoRR Hypothesis proposes that this Co-location is required for Redox Regulation.
Cyanobacteria and the evolution of photosynthesis
The biochemical capacity to use water as the source for electrons in photosynthesis evolved once, in a common ancestor of extant cyanobacteria. The geological record indicates that this transforming event took place early in Earth's history, at least 2450-2320 million years ago (Ma), and possibly much earlier.[citation needed] Available evidence from geobiological studies of Archean (>2500 Ma) sedimentary rocks indicates that life existed 3500 Ma, but the question of when oxygenic photosynthesis evolved is still unanswered. A clear paleontological window on cyanobacterial evolution opened about 2000 Ma, revealing an already-diverse biota of blue-greens. Cyanobacteria remained principal primary producers throughout the Proterozoic Eon (2500-543 Ma), in part because the redox structure of the oceans favored photoautotrophs capable of nitrogen fixation.[citation needed] Green algae joined blue-greens as major primary producers on continental shelves near the end of the Proterozoic, but only with the Mesozoic (251-65 Ma) radiations of dinoflagellates, coccolithophorids, and diatoms did primary production in marine shelf waters take modern form. Cyanobacteria remain critical to marine ecosystems as primary producers in oceanic gyres, as agents of biological nitrogen fixation, and, in modified form, as the plastids of marine algae.[28]
Discovery
Although some of the steps in photosynthesis are still not completely understood, the overall photosynthetic equation has been known since the 1800s.
Jan van Helmont began the research of the process in the mid-1600s when he carefully measured the mass of the soil used by a plant and the mass of the plant as it grew. After noticing that the soil mass changed very little, he hypothesized that the mass of the growing plant must come from the water, the only substance he added to the potted plant. His hypothesis was partially accurate—much of the gained mass also comes from carbon dioxide as well as water. However, this was a signaling point to the idea that the bulk of a plant's biomass comes from the inputs of photosynthesis, not the soil itself.
Joseph Priestley, a chemist and minister, discovered that when he isolated a volume of air under an inverted jar, and burned a candle in it, the candle would burn out very quickly, much before it ran out of wax. He further discovered that a mouse could similarly "injure" air. He then showed that the air that had been "injured" by the candle and the mouse could be restored by a plant.
In 1778, Jan Ingenhousz, court physician to the Austrian Empress, repeated Priestley's experiments. He discovered that it was the influence of sunlight on the plant that could cause it to rescue a mouse in a matter of hours.
In 1796, Jean Senebier, a Swiss pastor, botanist, and naturalist, demonstrated that green plants consume carbon dioxide and release oxygen under the influence of light. Soon afterwards, Nicolas-Théodore de Saussure showed that the increase in mass of the plant as it grows could not be due only to uptake of CO2, but also to the incorporation of water. Thus the basic reaction by which photosynthesis is used to produce food (such as glucose) was outlined.
Cornelis Van Niel made key discoveries explaining the chemistry of photosynthesis. By studying purple sulfur bacteria and green bacteria he was the first scientist to demonstrate that photosynthesis is a light-dependent redox reaction, in which hydrogen reduces carbon dioxide.
Robert Emerson discovered two light reactions by testing plant productivity using different wavelengths of light. With the red alone, the light reactions were suppressed. When blue and red were combined, the output was much more substantial. Thus, there were two photosystems, one aborbing up to 600 nm wavelengths, the other up to 700. The former is known as PSII, the latter is PSI. PSI contains only chlorophyll a, PSII contains primarily chlorophyll a with most of the available chlorophyll b, among other pigments.[29]
Further experiments to prove that the oxygen developed during the photosynthesis of green plants came from water, were performed by Robert Hill in 1937 and 1939. He showed that isolated chloroplasts give off oxygen in the presence of unnatural reducing agents like iron oxalate, ferricyanide or benzoquinone after exposure to light. The Hill reaction is as follows:
- 2 H2O + 2 A + (light, chloroplasts) → 2 AH2 + O2
where A is the electron acceptor. Therefore, in light the electron acceptor is reduced and oxygen is evolved. Cyt b6, now known as a plastoquinone, is one electron acceptor.
Samuel Ruben and Martin Kamen used radioactive isotopes to determine that the oxygen liberated in photosynthesis came from the water.
Melvin Calvin and Andrew Benson, along with James Bassham, elucidated the path of carbon assimilation (the photosynthetic carbon reduction cycle) in plants. The carbon reduction cycle is known as the Calvin cycle, which inappropriately ignores the contribution of Bassham and Benson. Many scientists refer to the cycle as the Calvin-Benson Cycle, Benson-Calvin, and some even call it the Calvin-Benson-Bassham (or CBB) Cycle.
A Nobel Prize winning scientist, Rudolph A. Marcus, was able to discover the function and significance of the electron transport chain.
Factors
There are three main factors affecting photosynthesis and several corollary factors. The three main are:
- Light irradiance and wavelength
- Carbon dioxide concentration
- Temperature.
Light intensity (irradiance), wavelength and temperature
In the early 1900s Frederick Frost Blackman along with Gabrielle Matthaei investigated the effects of light intensity (irradiance) and temperature on the rate of carbon assimilation.
- At constant temperature, the rate of carbon assimilation varies with irradiance, initially increasing as the irradiance increases. However at higher irradiance this relationship no longer holds and the rate of carbon assimilation reaches a plateau.
- At constant irradiance, the rate of carbon assimilation increases as the temperature is increased over a limited range. This effect is only seen at high irradiance levels. At low irradiance, increasing the temperature has little influence on the rate of carbon assimilation.
These two experiments illustrate vital points: firstly, from research it is known that photochemical reactions are not generally affected by temperature. However, these experiments clearly show that temperature affects the rate of carbon assimilation, so there must be two sets of reactions in the full process of carbon assimilation. These are of course the light-dependent 'photochemical' stage and the light-independent, temperature-dependent stage. Second, Blackman's experiments illustrate the concept of limiting factors. Another limiting factor is the wavelength of light. Cyanobacteria, which reside several meters underwater, cannot receive the correct wavelengths required to cause photoinduced charge separation in conventional photosynthetic pigments. To combat this problem, a series of proteins with different pigments surround the reaction center.This unit is called a phycobilisome.
Carbon dioxide levels and photorespiration
As carbon dioxide concentrations rise, the rate at which sugars are made by the light-independent reactions increases until limited by other factors. RuBisCO, the enzyme that captures carbon dioxide in the light-independent reactions, has a binding affinity for both carbon dioxide and oxygen. When the concentration of carbon dioxide is high, RuBisCO will fix carbon dioxide. However, if the oxygen concentration is high, RuBisCO will bind oxygen instead of carbon dioxide. This process, called photorespiration, uses energy, but does not make sugar.
RuBisCO oxygenase activity is disadvantageous to plants for several reasons:
- One product of oxygenase activity is phosphoglycolate (2 carbon) instead of 3-phosphoglycerate (3 carbon). Phosphoglycolate cannot be metabolized by the Calvin-Benson cycle and represents carbon lost from the cycle. A high oxygenase activity, therefore, drains the sugars that are required to recycle ribulose 5-bisphosphate and for the continuation of the Calvin-Benson cycle.
- Phosphoglycolate is quickly metabolized to glycolate that is toxic to a plant at a high concentration; it inhibits photosynthesis.
- Salvaging glycolate is an energetically expensive process that uses the glycolate pathway and only 75% of the carbon is returned to the Calvin-Benson cycle as 3-phosphoglycerate.
-
- A highly-simplified summary is:
-
-
- 2 glycolate + ATP → 3-phosphoglycerate + carbon dioxide + ADP +NH3
-
The salvaging pathway for the products of RuBisCO oxygenase activity is more commonly known as photorespiration, since it is characterized by light-dependent oxygen consumption and the release of carbon dioxide.
See also
Footnotes
α. ^ The word photosynthesis comes from the Greek φώτο- (photo-), "light," and σύνθεσις (synthesis), "placing with."
β. ^ The exceptions are chemoautotrophs that live in rocks or around deep sea hydrothermal vents.
References
|
|
This article needs additional citations for verification. Please help improve this article by adding reliable references (ideally, using inline citations). Unsourced material may be challenged and removed. (August 2007) |
- ^ Smith, A. L. (1997). Oxford dictionary of biochemistry and molecular biology. Oxford [Oxfordshire]: Oxford University Press. pp. 508. ISBN 0-19-854768-4. "Photosynthesis - the synthesis by organisms of organic chemical compounds, esp. carbohydrates, from carbon dioxide using energy obtained from light rather than the oxidation of chemical compounds."
- ^ a b D.A. Bryant & N.-U. Frigaard (November 2006). "Prokaryotic photosynthesis and phototrophy illuminated". Trends Microbiol 14 (11): 488. doi:.
- ^ Nealson KH, Conrad PG (December 1999). "Life: past, present and future". Philos. Trans. R. Soc. Lond., B, Biol. Sci. 354 (1392): 1923–39. doi:. PMID 10670014. PMC: 1692713. http://journals.royalsociety.org/content/7r10hqn3rp1g1vag/.
- ^ "World Consumption of Primary Energy by Energy Type and Selected Country Groups , 1980-2004" (XLS). Energy Information Administration. July 31, 2006. http://www.eia.doe.gov/pub/international/iealf/table18.xls.
- ^ Field CB, Behrenfeld MJ, Randerson JT, Falkowski P (July 1998). "Primary production of the biosphere: integrating terrestrial and oceanic components". Science (journal) 281 (5374): 237–40. doi:. PMID 9657713.
- ^ Olson JM (May 2006). "Photosynthesis in the Archean era". Photosyn. Res. 88 (2): 109–17. doi:. PMID 16453059.
- ^ Buick R (August 2008). "When did oxygenic photosynthesis evolve?". Philos. Trans. R. Soc. Lond., B, Biol. Sci. 363 (1504): 2731–43. doi:. PMID 18468984.
- ^ Gould SB, Waller RF, McFadden GI (2008). "Plastid evolution". Annu Rev Plant Biol 59: 491–517. doi:. PMID 18315522.
- ^ Anaerobic Photosynthesis, Chemical & Engineering News, 86, 33, 18 Aug. 2008, p. 36
- ^ Kulp TR, Hoeft SE, Asao M, Madigan MT, Hollibaugh JT, Fisher JC, Stolz JF, Culbertson CW, Miller LG, Oremland RS (August 2008). "Arsenic(III) fuels anoxygenic photosynthesis in hot spring biofilms from Mono Lake, California". Science (journal) 321 (5891): 967–70. doi:. PMID 18703741.
- ^ Tavano CL, Donohue TJ (December 2006). "Development of the bacterial photosynthetic apparatus". Curr. Opin. Microbiol. 9 (6): 625–31. doi:. PMID 17055774.
- ^ a b Mullineaux CW (1999). "The thylakoid membranes of cyanobacteria: structure, dynamics and function". Australian Journal of Plant Physiology 26 (7): 671–677. doi:.
- ^ Sener MK, Olsen JD, Hunter CN, Schulten K (October 2007). "Atomic-level structural and functional model of a bacterial photosynthetic membrane vesicle". Proc. Natl. Acad. Sci. U.S.A. 104 (40): 15723–8. doi:. PMID 17895378. PMC: 2000399. http://www.pnas.org/cgi/pmidlookup?view=long&pmid=17895378.
- ^ Campbell, Neil A.; Brad Williamson; Robin J. Heyden (2006). Biology: Exploring Life. Boston, Massachusetts: Pearson Prentice Hall. ISBN 0-13-250882-6. http://www.phschool.com/el_marketing.html.
- ^ a b Raven, Peter H.; Ray F. Evert, Susan E. Eichhorn (2005). Biology of Plants, 7th Edition. New York: W.H. Freeman and Company Publishers. pp. 124–127. ISBN 0-7167-1007-2.
- ^ "Yachandra Group Home page". http://www.lbl.gov/~vkyachan/index.html.
- ^ Pushkar Y, Yano J, Sauer K, Boussac A, Yachandra VK (February 2008). "Structural changes in the Mn4Ca cluster and the mechanism of photosynthetic water splitting". Proc. Natl. Acad. Sci. U.S.A. 105 (6): 1879–84. doi:. PMID 18250316.
- ^ L. Taiz, E. Zeiger (2006). Plant Physiology (4 ed.). Sinauer Associates. ISBN 978-0878938568.
- ^ Miyamoto K. "Chapter 1 - Biological energy production". Renewable biological systems for alternative sustainable energy production (FAO Agricultural Services Bulletin - 128). Food and Agriculture Organization of the United Nations. http://www.fao.org/docrep/w7241e/w7241e05.htm#1.2.1%20photosynthetic%20efficiency. Retrieved on 2009-01-04.
- ^ New Scientist, 19 Aug., 2006
- ^ Venn AA, Loram JE, Douglas AE (2008). "Photosynthetic symbioses in animals". J. Exp. Bot. 59 (5): 1069–80. doi:. PMID 18267943. http://jxb.oxfordjournals.org/cgi/content/full/59/5/1069.
- ^ Rumpho ME, Summer EJ, Manhart JR (May 2000). "Solar-powered sea slugs. Mollusc/algal chloroplast symbiosis". Plant Physiol. 123 (1): 29–38. doi:. PMID 10806222.
- ^ Muscatine L, Greene RW (1973). "Chloroplasts and algae as symbionts in molluscs". Int. Rev. Cytol. 36: 137–69. PMID 4587388.
- ^ Rumpho ME, Worful JM, Lee J, et al (November 2008). "From the Cover: Horizontal gene transfer of the algal nuclear gene psbO to the photosynthetic sea slug Elysia chlorotica". Proc. Natl. Acad. Sci. U.S.A. 105 (46): 17867–17871. doi:. PMID 19004808.
- ^ Douglas SE (December 1998). "Plastid evolution: origins, diversity, trends". Curr. Opin. Genet. Dev. 8 (6): 655–61. PMID 9914199. http://linkinghub.elsevier.com/retrieve/pii/S0959-437X(98)80033-6.
- ^ Reyes-Prieto A, Weber AP, Bhattacharya D (2007). "The origin and establishment of the plastid in algae and plants". Annu. Rev. Genet. 41: 147–68. doi:. PMID 17600460.
- ^ Raven JA, Allen JF (2003). "Genomics and chloroplast evolution: what did cyanobacteria do for plants?". Genome Biol. 4 (3): 209. doi:. PMID 12620099. PMC: 153454. http://genomebiology.com/1465-6906/4/209.
- ^ Herrero A and Flores E (editor). (2008). The Cyanobacteria: Molecular Biology, Genomics and Evolution (1st ed. ed.). Caister Academic Press. ISBN 978-1-904455-15-8.
- ^ McGraw Hill Encyclopedia of Science & Technology (Mcgraw Hill Encyclopedia of Science and Technology). McGraw-Hill Professional. 2007. pp. vol 13 p. 470. ISBN 0-07-144143-3.
Further reading
- Asimov, Isaac (1968). Photosynthesis. New York, London: Basic Books, Inc.. ISBN 0-465-05703-9.
- Bidlack JE; Stern KR, Jansky S (2003). Introductory plant biology. New York: McGraw-Hill. ISBN 0-07-290941-2.
- Blankenship RE (2008). Molecular Mechanisms of Photosynthesis (2nd ed.). John Wiley & Sons Inc. ISBN 0-470-71451-4.
- Govindjee (1975). Bioenergetics of photosynthesis. Boston: Academic Press. ISBN 0-12-294350-3.
- Govindjee Beatty JT,Gest H, Allen JF (2006). Discoveries in Photosynthesis. Advances in Photosynthesis and Respiration. 20. Berlin: Springer. ISBN 1-4020-3323-0.
- Gregory RL (1971). Biochemistry of photosynthesis. New York: Wiley-Interscience. ISBN 0-471-32675-5.
- Rabinowitch E, Govindjee (1969). Photosynthesis. London: J. Wiley. ISBN 0-471-70424-5.
- Reece, J, Campbell, N (2005). Biology. San Francisco: Pearson, Benjamin Cummings. ISBN 0-8053-7146-X.
External links
| Wikimedia Commons has media related to: Photosynthesis |
- A collection of photosynthesis pages for all levels from a renowned expert (Govindjee)
- UC Berkeley video lecture on Photosynthesis
- In depth, advanced treatment of photosynthesis, also from Govindjee
- Science Aid: Photosynthesis Article appropriate for high school science
- Liverpool John Moores University, Dr.David Wilkinson
- Metabolism, Cellular Respiration and Photosynthesis - The Virtual Library of Biochemistry and Cell Biology
- Overall examination of Photosynthesis at an intermediate level
- Overall Energetics of Photosynthesis
- Photosynthesis Discovery Milestones - experiments and background
|
||||||||||||||||||||||||||


